RETRACTED: Pyrophosphate modulates plant stress responses via SUMOylation
- Zaida Andrés

- Melanie Krebs
- Fabian Fink
- Katarzyna Drzewicka
- Nicolas Stankovic-Valentin
- Shoji Segami
- Sebastian Schuck
- Michael Büttner
- Rüdiger Hell
- Masayoshi Maeshima
- Frauke Melchior
- Karin Schumacher
- Heidelberg University, Germany
- Center for Molecular Biology of Heidelberg University (ZMBH) and DKFZ - ZMBH Alliance, Germany
- Nagoya University, Japan
Abstract
Pyrophosphate (PPi), a byproduct of macromolecule biosynthesis is maintained at low levels by soluble inorganic pyrophosphatases (sPPase) found in all eukaryotes. In plants, H+-pumping pyrophosphatases (H+-PPase) convert the substantial energy present in PPi into an electrochemical gradient. We show here, that both cold- and heat stress sensitivity of fugu5 mutants lacking the major H+-PPase isoform AVP1 is correlated with reduced SUMOylation. In addition, we show that increased PPi concentrations interfere with SUMOylation in yeast and we provide evidence that SUMO activating E1-enzymes are inhibited by micromolar concentrations of PPi in a non-competitive manner. Taken together, our results do not only provide a mechanistic explanation for the beneficial effects of AVP1 overexpression in plants but they also highlight PPi as an important integrator of metabolism and stress tolerance.
https://doi.org/10.7554/eLife.44213.001Introduction
Reshaping of metabolic networks under stress conditions enables the synthesis of protective compounds while metabolic homeostasis needs to be maintained. In about 200 metabolic reactions ATP is not used as a phosphorylating but as an adenylating reagent leading to the release of inorganic pyrophosphate (PPi). Most prominently, the biosynthesis of many macromolecules including DNA, RNA, proteins and polysaccharides releases large amounts of PPi (Ferjani et al., 2014; Heinonen, 2001). Given the substantial free energy of PPi, the efficient biosynthesis of macromolecules requires that PPi is immediately destroyed to prevent the respective back-reactions (Kornberg, 1962). In all eukaryotes PPi is hydrolysed by soluble inorganic pyrophosphatases (sPPase; EC 3.6.1.1) in a highly exergonic reaction. Loss of sPPase function causes lethality in yeast (Lundin et al., 1991) and C. elegans (Ko et al., 2007) presumably due to accumulation of PPi inhibiting the biosynthesis of macromolecules. Arabidopsis encodes six sPPase-paralogs (PPa1-PPa6) of which only PPa6 is localized in plastids whereas all others are cytosolic (Gutiérrez-Luna et al., 2016; Segami et al., 2018). However, their PPase activity is rather low and even the loss of the four ubiquitously expressed isoforms does not cause visible phenotypic alterations (Segami et al., 2018). In contrast, expression of E. coli sPPase severely affects plant growth via alterations in carbon partitioning between source and sink organs caused by the inhibition of several plant enzymes involved in carbohydrate metabolism that use PPi as an energy source (Geigenberger et al., 1998; Sonnewald, 1992). Importantly, in addition to soluble PPases, plants contain membrane-bound proton-pumping pyrophosphatases (H+-PPase) at the tonoplast and in the Golgi that convert the energy otherwise released as heat into a proton-gradient (Maeshima, 2000; Segami et al., 2010). Mutants lacking the tonoplast H+-PPase AVP1 (Arabidopsis vacuolar H+-PPase) were identified based on their compensatory cell enlargement phenotype and were thus named fugu5 (after the japanese term for the pufferfish; Ferjani et al., 2011). The fact that the fugu5 phenotype could be rescued either by growth in the presence of exogenous sucrose or the expression of the yeast sPPase IPP1 showed clearly that altered PPi levels and not reduced H+-pumping are causative (Asaoka et al., 2016; Ferjani et al., 2011). Indeed, vacuolar pH is only mildly affected in fugu5 mutants indicating that the H+-pumping ATPase (V-ATPase) present at the tonoplast is largely sufficient for vacuolar acidification (Ferjani et al., 2011; Kriegel et al., 2015). However, loss of both vacuolar proton-pumps leads to a much more severe phenotype and defect in vacuolar acidification than loss of the tonoplast V-ATPase alone (Kriegel et al., 2015). It has indeed been discussed that AVP1 serves as a backup system for the V-ATPase in particular under ATP-limiting conditions like anoxia or cold stress (Maeshima, 2000). During cold acclimation plants accumulate cryoprotectants including sugars in their vacuoles and activity of both proton-pumps is upregulated leading to improved freezing tolerance (Schulze et al., 2012; Thomashow, 1999). Overexpression of AVP1 has been shown to cause increased plant growth under various abiotic stress conditions including salinity, drought and phosphate starvation but the underlying mechanism remained unclear (Gaxiola et al., 2012; Park et al., 2005; Schilling et al., 2017). Attachment of the small ubiquitin-related modifier SUMO to substrate proteins plays a central role in the response to a broad set of stress responses including the ones affected by AVP1 overexpression (Castro et al., 2012). Modification of target proteins by SUMO-conjugation proceeds via a three-step mechanism. First the SUMO moiety is adenylated and then bound via a high-energy thioester linkage to the heterodimeric SUMO-activating enzyme (E1) leading to the release of PPi. Next, the activated SUMO is transferred to the SUMO-conjugating enzyme E2 and finally, assisted by SUMO-protein ligase (E3), donated to a large set of substrate proteins (Flotho and Melchior, 2013; Johnson, 2004). In Arabidopsis, the key transcriptional regulator of the cold response Inducer of CBF expression 1 (ICE1) as well as the Heat shock factor A2 (HSFA2) have been shown to be positively regulated by SUMOylation (Cohen-Peer et al., 2010; Miura et al., 2007). In this study, we report that AVP1 contributes to both cold acclimation and heat tolerance and we show that the rapid increase in SUMOylation common to both stress responses is missing in the absence of AVP1. Furthermore, we provide evidence that accumulation of PPi in plants, yeast and mammals inhibits the SUMO E1 activating enzyme in turn affecting the fate, localization or function of a large number of proteins during cellular stress responses. Our results provide a mechanistic explanation for the beneficial effects of AVP1 overexpression in plants and highlight PPi as an important integrator of metabolism and stress tolerance.
Results
Lack of V-PPase activity impairs cold acclimation
We have shown previously that upregulation of ATP-hydrolysis by the V-ATPase during cold acclimation depends on the presence and the activity of the V-PPase (Kriegel et al., 2015). To complete the data-set for vacuolar proton-pump activity during cold acclimation, we performed parallel measurements of ATP- and PPi- hydrolysis, H+-pumping as well as cell sap pH in wild-type (Col-0), the fugu5-1 mutant and a UBQ:AVP1 overexpression line. Both ATP- and PPi-dependent proton-pumping are increased in wild-type (wt) and UBQ:AVP1 during cold-acclimation (Figure 1—figure supplement 1A, B). As expected PPi-dependent proton-pumping was undetectable in fugu5-1, but ATP-dependent proton-pumping was also reduced in fugu5-1 compared to wt and increased only marginally upon cold-acclimation (Figure 1—figure supplement 1B). As a consequence of cold-induced proton-pump stimulation, vacuolar pH drops by 0.1 pH-units in wt and UBQ:AVP1 but not in fugu5-1 (Figure 1—figure supplement 1C).
Whereas the seedling phenotype of fugu5-1 is rescued by expression of the yeast soluble PPase IPP1 under control of the AVP1-promoter during the seedling stage (Ferjani et al., 2011), the adult growth phenotype of plants grown in short day was not rescued (Figure 1—figure supplement 2A).
We thus asked if constitutive overexpression of an Arabidopsis sPPase would be able to fully rescue the fugu5 phenotype. Among four broadly expressed Arabidopsis sPPase genes we chose PPa5 and expressed its coding sequence fused to GFP under the control of the UBQ10-promoter in both wt and fugu5-1. In the resulting transgenic lines PPa5-GFP was located in the cytosol as well as in the nucleus (Figure 1—figure supplement 2B) and the seedling (Figure 1—figure supplement 2C) as well as the adult phenotype (Figure 1—figure supplement 2D+E) of fugu5-1 were fully rescued. For further analysis, we chose two lines in the wt and in the fugu5-1 background with comparable increased total soluble pyrophosphatase activity (Figure 1—figure supplement 2F). Interestingly, PPa5-GFP accumulated to much higher levels in fugu5-1 than in wt (Figure 1—figure supplement 2G) indicating that the fugu5-1 mutation influences either protein stability or transcript levels of PPa5-GFP.
We next asked if cold-acclimation is affected in fugu5-1 and if so, whether this could be rescued by overexpression of a soluble pyrophosphatase. Both survival rate and ion release as a measure of freezing tolerance was comparable in all genotypes exposed to freezing without prior cold acclimation (Figure 1A + B). Cold acclimation via exposure to 4°C for 4 days significantly improved freezing tolerance in wt to a much higher extent than in fugu5-1 plants and expression of PPa5 fully rescued the hypersensitivity to cold (Figure 1C). Conversely, mutants lacking one or several sPPases (Segami et al., 2018) are less freezing tolerant prior to cold exposure but respond like wt after cold-acclimation (Figure 1—figure supplement 3).
Figure 1 with 3 supplements see all
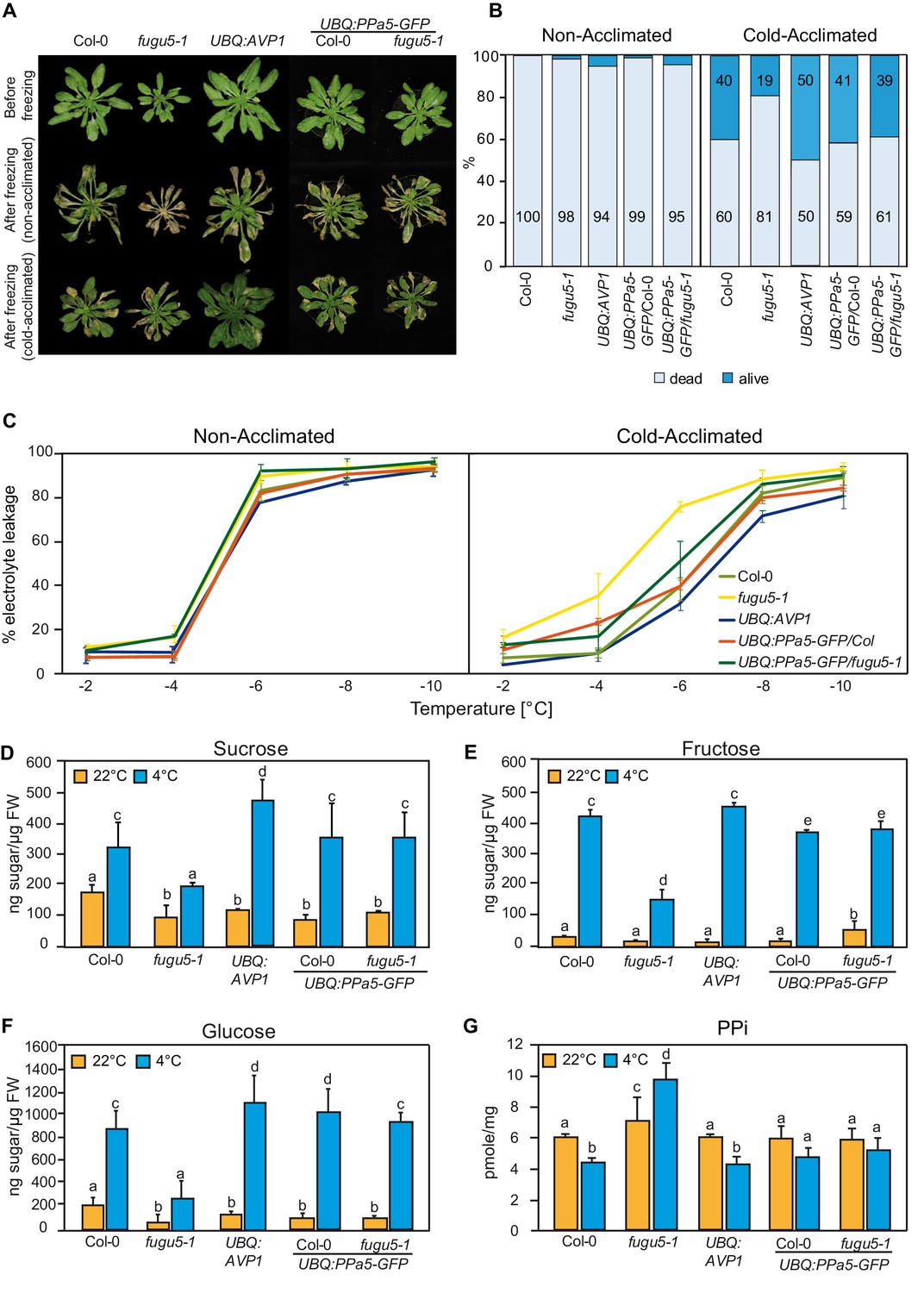
PPi hydrolysis is required to rescue the freezing sensitive phenotype of fugu5-1.
(A) and (B) Freezing tolerance assay. Col-0, fugu5-1, UBQ:AVP1 and UBQ:PPa5-GFP in Col-0 and fugu5-1 backgrounds were grown for 6 weeks at 22°C and were then moved to 4°C for cold-acclimation, or kept at 22°C for 4 days. Afterwards, plants were subjected to a 5 hr freezing temperature regime (0 to −10°C). After thawing at 4°C overnight, plants were moved back to 22°C. (A) Images were taken before cold-acclimation and one week after the freezing treatment. (B) Quantification of dead and alive leaves was done one week after the freezing treatment with n ≥ 75 leaves from 5 individual plants. 3 independent experiments were performed. Cold-acclimated UBQ:AVP1 and fugu5-1 are significantly different compared to the other genotypes (Student’s t-test p<0.05). (C) Electrolyte leakage assay of Col-0, fugu5-1, UBQ:AVP1 and UBQ:PPa5-GFP in Col-0 and fugu5-1 backgrounds was performed on leaf material of acclimated and non-acclimated plants at indicated freezing temperatures. Error bars represent SD of the mean of n = 3 biological replicates. (D–G) Sugar and PPi measurements were done from extracts of acclimated (4°C) and non-acclimated (22°C) 6-week-old rosette leaves. Error bars show SD of the mean with n = 3 samples of one representative experiments. 3 biological replicates were performed. Significant differences are indicated by different letters (Two-way ANOVA followed by Tukey’s test, p<0.05).
-
Figure 1—source data 1
(B) Survival of the 6 weeks old rosette leaves upon freezing.
(C) Electrolyte leakage analysis of the cold-acclimated and non-acclimated 6-weeks old rosette leaves upon exposure to freezing temperature. (D-G) Sugar and PPi measurements of the 6-weeks old rosette leaves.
- https://doi.org/10.7554/eLife.44213.008
Accumulation of soluble sugars during exposure to low temperatures contributes to freezing tolerance and could be directly affected by PPi-accumulation (Ferjani et al., 2018). We thus next compared the accumulation of glucose, fructose and sucrose and found that cold-induced sugar accumulation is indeed strongly reduced in the fugu5-1 mutant but restored by UBQ:PPa5-GFP (Figure 1D–1F) suggesting that accumulation of PPi and not a lack of H+-pumping is responsible for the impaired cold acclimation in fugu5-1. In agreement with this hypothesis, we found that PPi levels are reduced in the wt during cold-acclimation whereas they increase in fugu5-1 resulting in 2-fold higher levels compared to wt after cold acclimation (Figure 1G).
PPi controls cold-acclimation potentially via SUMOylation of ICE1
Low temperature triggers the expression of the CBF (C-repeat binding factor) family of transcription factors, which in turn activate downstream genes that confer chilling and freezing tolerance (Chinnusamy et al., 2007). We used qRT-PCR to profile the expression of members of the PPa-gene family over 24 hr after exposure to low temperature (4°C) and found that PPa1 and PPa4 are rapidly induced after cold exposure whereas transcripts of PPa2 and PPa5 accumulated at later time points (Figure 2A). Upregulation of sPPase genes suggests that PPi-levels are actively controlled during the early cold acclimation response. In fugu5, cold-induced upregulation of PPa1, PPa4 and PPa5 is reduced (Figure 2A) and we thus next compared the expression levels of the core transcriptional regulators CBF1-3 as well as the three downstream response genes COR15A, COR78 and GolS3. Whereas expression of CBF2 was nearly unaffected, CBF1 and in particular CBF3 induction was found to be strongly reduced in the fugu5-1 mutant (Figure 2B). Similarly, induction of all three target genes was found to be strongly reduced throughout the cold response (Figure 2C). Cold-induction was restored when PPa5 was constitutively expressed in the fugu5-1 background (Figure 2 B + C) indicating that the observed changes in gene expression are caused by reduced PPi-hydrolysis and not by reduced H+-pumping.
Figure 2

sPPase expression is induced upon cold exposure to control PPi that affects expression of CBFs and CBF target genes.
(A) qRT-PCR for the analysis of expression of PPa 1,2,4,5 and AVP1 in Col-0 and PPa 1,2,4 and 5 in fugu5-1. (B-C) Measurement of expression of CBF, COR and GolS3 genes in Col-0, fugu5-1 and UBQ:PPa5-GFP/fugu5-1 by qRT-PCR. (A-C) and () Plants were grown for six weeks under short-day conditions at 22°C. Afterwards, they were exposed to 4°C for indicated time periods. Whole rosettes were used for total RNA extraction. Actin2 expression was used for normalization. Error bars represent SD of the mean of n = 3 biological replicates. Data analysis was performed using the ΔΔCt method.
-
Figure 2—source data 1
(A) Relative expression of the vacuolar and soluble pyrophosphatases in wt and the relative expression of the soluble pyrophosphtases in fugu5-1 exposed to 4°C for different hours.
(B-C) Relative expression of the cold regulated genes in wt, fugu5-1 and UBQ:PPa5-GFP/fugu5-1 upon exposure to 4°C for different hours.
- https://doi.org/10.7554/eLife.44213.010
The fast transcriptional response to cold is initiated by ICE1 (Inducer of CBF expression 1), a direct activator that is negatively regulated by ubiquitination-mediated proteolysis and positively regulated by SUMOylation (Dong et al., 2006; Miura et al., 2007; Figure 3A). Using a specific antibody to detect ICE1 in total seedling protein extracted in the presence of NEM to inhibit deSUMOylation, we observed two bands in wild-type that are both absent in ice1-2 indicating that they might correspond to a non-modified (100kD) and modified (130kD) dimer of ICE1. The ICE1 monomer (50kD) was only observed when proteins were extracted in the presence of DTT and without NEM (Figure 3—figure supplement 1).
Figure 3 with 1 supplement see all
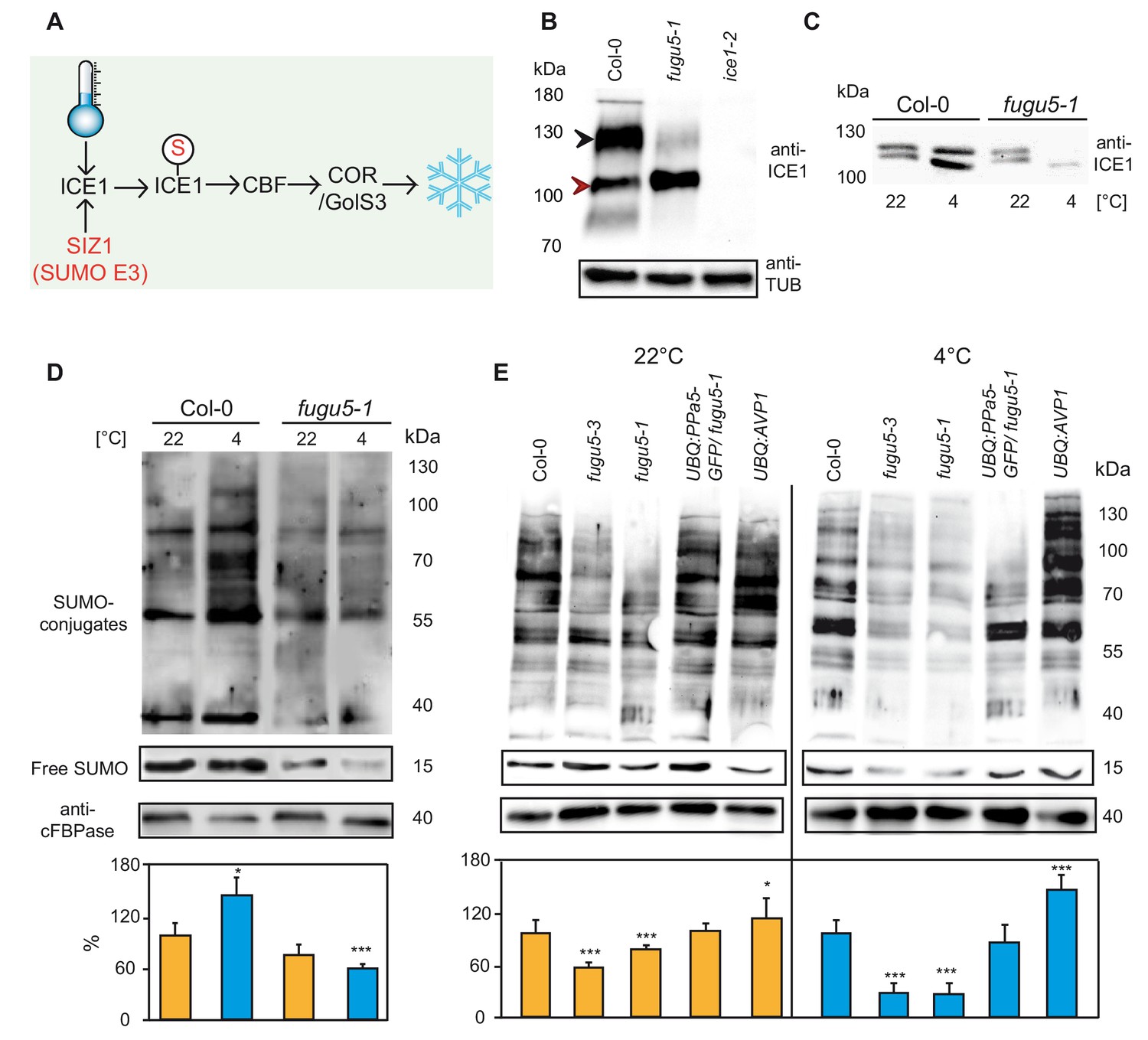
Modification of ICE1 and general SUMOylation upon cold exposure are inhibited in fugu5.
(A) Cold acclimation induces ICE1 SUMOylation which then activates CBFs and leads to the expression of downstream targets for the establishment of freezing tolerance. (B) Determination of the amount of ICE1 in Col-0, fugu5-1 and ice1-2 seedlings. Amount of TUBULIN was detected as loading control. Plants grown in long day conditions for 3 weeks. Black arrow indicates modified ICE1 dimer whereas red arrow indicates unmodified ICE1 dimer. (C) Comparison of the amount of the ICE1 under normal conditions (22°C) and after cold treatment (4°C, 3 hr) in Col-0 and fugu5-1 seedlings. 10-days-old liquid grown seedlings were used for total protein extraction. Anti-ICE1 was used as primary antibody. (D) Western blots comparing SUMOylation levels of Col-0 and fugu5-1 under normal conditions (22°C) and after cold treatment (4°C, 3 hr). (E) Western blots demonstrating the total SUMOylation in Col-0, fugu5-1, fugu5-3, UBQ:AVP1 and UBQ:PPa5-GFP under normal conditions (22°C) and after cold treatment (4°C, 3 hr). (D) and (E) 10 days old liquid grown seedlings were used for total protein extraction. Anti-SUMO1/2 was used as primary antibody. Whole lanes were measured for the calculation of protein amounts using ImageJ. cFBPase detection was used for normalization. Error bars represent SD of n ≥ 2 biological replicates. Asterisk indicates significant difference compared to Col-0 (Student’s t test; *p<0.05, ***p<0.001).
-
Figure 3—source data 1
(D-E) Comparison of the amount of the total SUMOylation with and without cold treatment.
- https://doi.org/10.7554/eLife.44213.013
In fugu5-1 the modified dimer was barely detectable indicating that either ubiquitination or SUMOylation of ICE1 are affected (Figure 3B). We next compared levels of ICE1 during cold acclimation and found that ICE1 accumulated after exposure to 4°C for 3 hr in wt but was strongly reduced in fugu5-1 (Figure 3C). We thus asked if overall cold-induced SUMOylation was affected in fugu5. Whereas cold exposure let to a rapid and massive accumulation SUMO1/2 conjugates in the wild-type, this response was absent in fugu5-1 (Figure 3D). Quantification of SUMO levels in two independent fugu5-alleles showed that it is already reduced to 60% of wt in plants grown at 22°C and levels drop to 20% of wt after incubation at 4°C for 3 hr (Figure 3E). SUMOylation is restored to wild-type levels in UBQ:PPa5-GFP and slightly enhanced in UBQ:AVP1 plants indicating that low PPi levels are critical for efficient SUMOylation (Figure 3D).
PPi inhibits heat-stress induced SUMOylation in both plants and yeast
Rapid and reversible accumulation of SUMO conjugates does not only occur during cold stress but also during heat stress (Miller et al., 2010; Rytz et al., 2018) and accumulation of PPi should thus also inhibit the heat stress response. Indeed, survival rate of fugu5 seedlings was strongly reduced by exposure to 40°C for 30 min but restored in the UBQ:PPa5-GFP complementation line. Of note, the survival rate of the UBQ:AVP1 overexpression line was increased compared to the wt (Figure 4A and B). We therefore analysed next if heat induced SUMO conjugate accumulation was affected. Exposure to 40°C for 30 min led to accumulation of SUMO conjugates in the wt, whereas the response was strongly reduced in fugu5-1 (Figure 4C). Consistently, a reduction of SUMO levels after heat stress was also observed for fugu5-3 but was restored to wt levels in UBQ:PPa5-GFP and UBQ:AVP1 plants (Figure 4D).
Figure 4
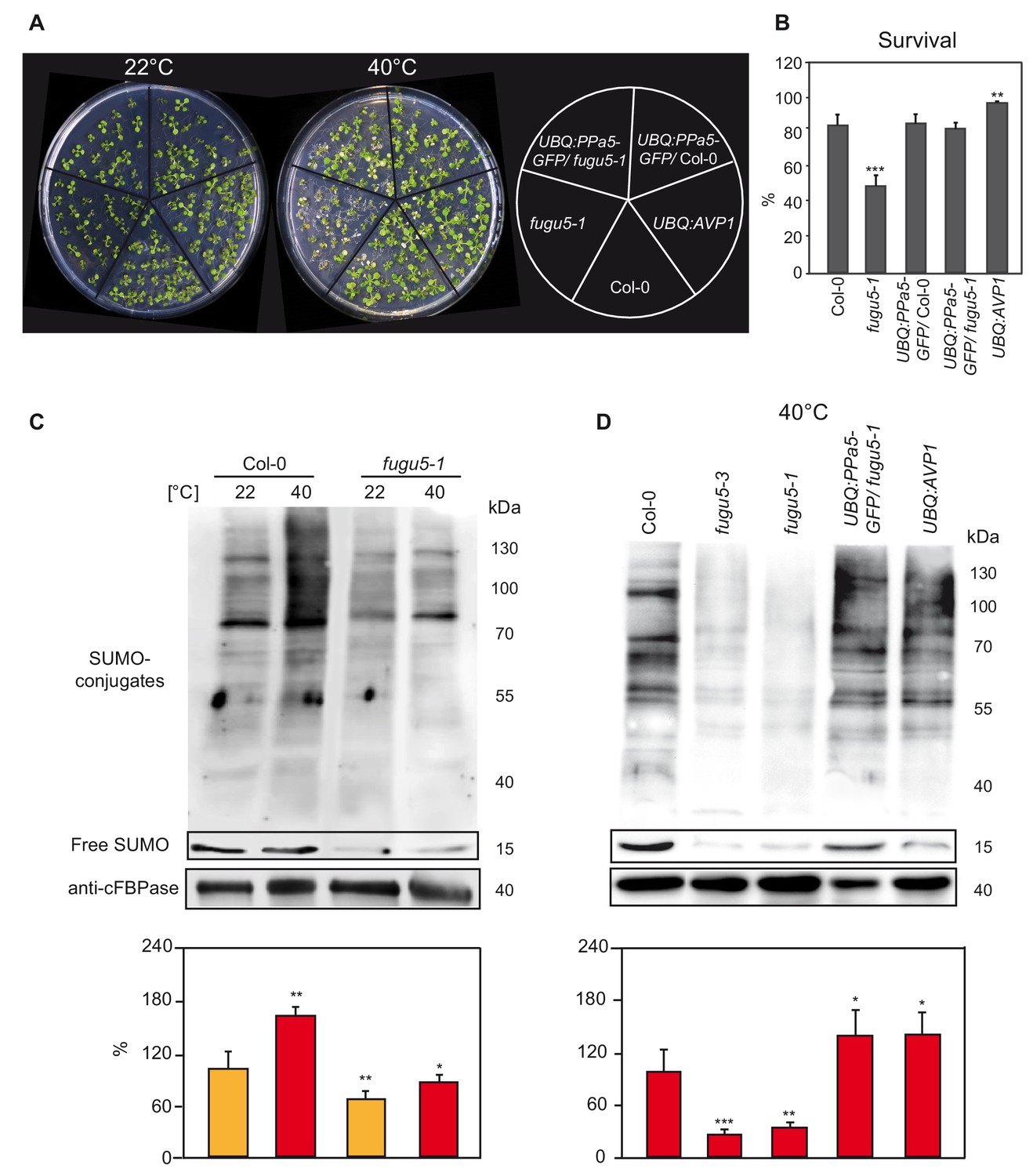
Heat shock-induced SUMOylation is also reduced in fugu5.
(A) Phenotypic analysis of 10-days-old Col-0, fugu5-1, UBQ:PPa5-GFP in Col-0 and fugu5-1 backgrounds and UBQ:AVP1 seedlings analysis before and after heat. Representative pictures of seedlings before and 4 days after completion of heat shock treatment (40°C, 4 hours) are depicted. (B) Seedling survival was determined 4 days after the heat shock. Alive and dead seedlings were counted and survival is shown as the percentage of the living seedlings. Error bars show SD of the mean with n ≥ 24 samples of one representative experiment. Two biological experiments were performed. Asterisk indicates significant difference compared to Col-0 (Student’s t test; **p<0.01, ***p<0.001). (C) SUMOylation levels of Col-0 and V-PPase mutant fugu5-1 were analysed with western blot under normal conditions (22°C) and after heat shock treatment (40°C, 30 min). (D) Measurement of the SUMO amount of Col-0, V-PPase mutants, UBQ:PPa5-GFP/fugu5-1 and UBQ:AVP1 seedlings after heat shock treatment (40°C, 30 min). (C) and (D) 10-days-old liquid grown seedlings are used for total protein extraction. Anti-SUMO1/2 (Agrisera) was used as primary antibody. Whole lanes were measured for the calculation of protein amounts using ImageJ. cFBPase detection was used for normalization. Error bars represent SD of n = 2 biological replicates. Asterisk indicates significant difference compared to Col-0 (Student’s t test; *p<0.05, **p<0.01, ***p<0.001).
-
Figure 4—source data 1
(B) Survival measurement of the 10 days old seedlings upon heat shock.
(C-D) Comparison of the amount of the total SUMOylation with and without heat shock treatment.
- https://doi.org/10.7554/eLife.44213.015
SUMO plays an important role in stress responses across all eukaryotes (Enserink, 2015; Hannich et al., 2005). Therefore, we asked whether PPi accumulation has a comparable effect in the yeast S. cerevisiae. We employed a strain in which the sole and essential sPPase IPP1 is expressed under the control of the GAL1 promoter (Serrano-Bueno et al., 2013) so that switching the carbon source from galactose to glucose led to a depletion of IPP1 (Figure 5A) after 6 hr that was almost complete after 15 hr (Figure 5A). When wt yeast was subjected to heat stress (40°C, 1 hr), SUMOylation increased by a factor of two (Figure 5B). Heat-induced SUMOylation was strongly diminished by depletion of IPP1, (Figure 5C) indicating that inhibition of SUMOylation by PPi is not limited to plants.
Figure 5
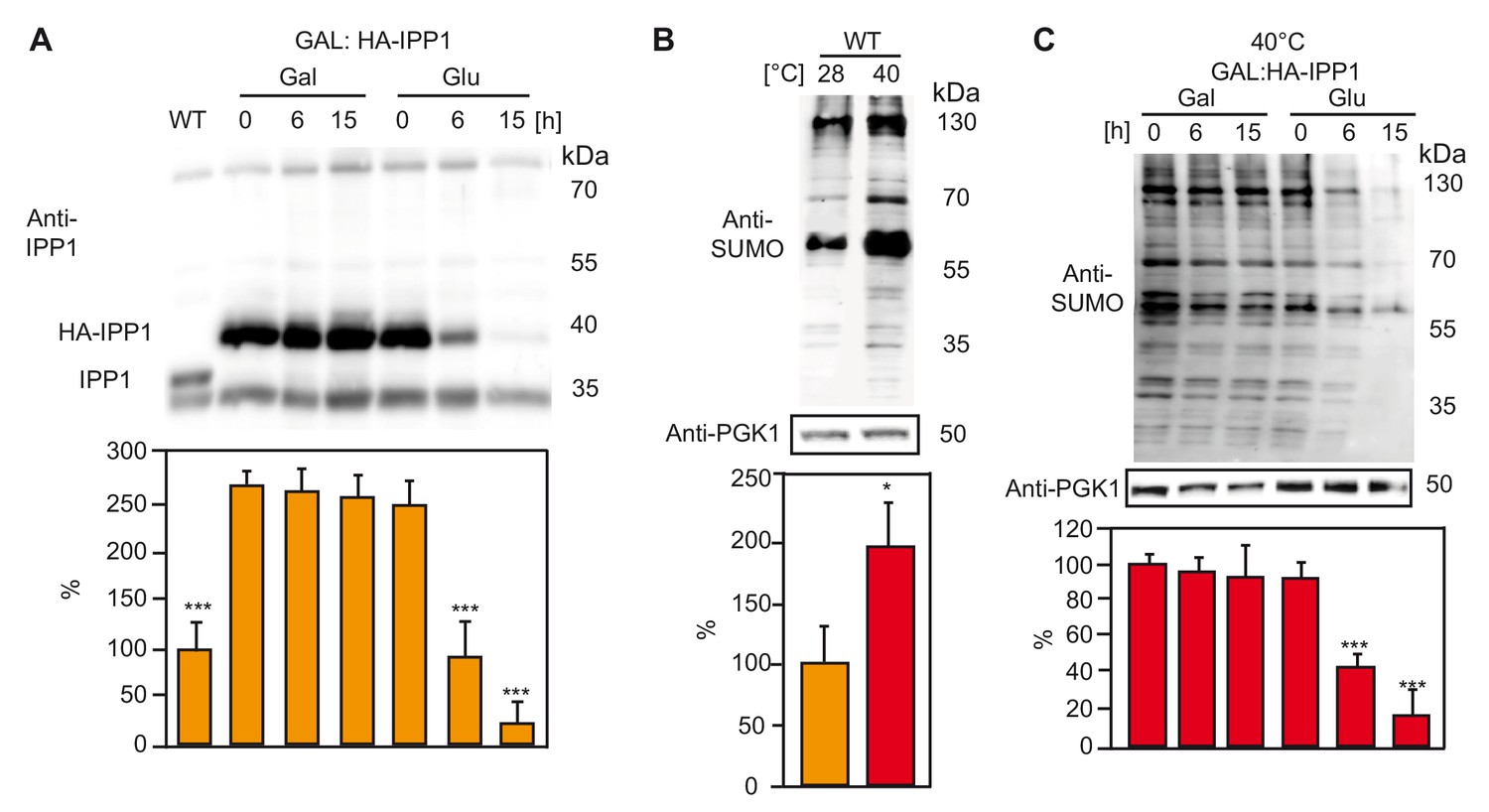
Increased PPi levels interfere with SUMOylation in yeast.
(A) Amount of the soluble pyrophosphatase protein in a conditional Ipp1 mutant of S. cerevisiae (GAL:HA-IPP1). Asterisk indicates significant difference compared to GAL:HA-IPP1 in galactose time point 0 hr (Student’s t test; ***p<0.001). Wt strain (W303) was used as a control. (B) Amount of total SUMOylation in W303 determined before and after heat stress. Asterisk indicates significant difference (Student’s t test; *p<0.05) (C) Measurement of total SUMO protein in conditional IPP1 mutant of S. cerevisiae (GAL:HA-IPP1). Asterisk indicates significant difference (Student’s t test; ***p<0.001). (A–C) Yeast is grown in synthetic complete medium supplemented with galactose at 28°C. After growing until OD600 0.5, part of the Ipp1 conditional mutant is switched to glucose supplemented medium to suppress the promoter and samples are collected at the indicated time points. For the heat treatment, cultures were switched to 40°C incubator for 1 hr. Error bars represent SD of n ≥ 2 biological replicates.
-
Figure 5—source data 1
(A) Amount of IPP1 in different carbon supplies over time.
(B) Comparison of the total SUMOylation of wt yeast at 28 and 40°C. (C) Comparison of the total SUMOylation at 40°C, in Ipp1 conditional mutant.
- https://doi.org/10.7554/eLife.44213.017
What is the mechanistic link between PPi accumulation and SUMOylation?
Conjugation of SUMO to target proteins is initiated by E1 enzymes through adenylation, a reaction that releases PPi and could thus be inhibited by increased cytosolic PPi levels (Haas et al., 1982; Lois and Lima, 2005). To test the direct effect of PPi on SUMOylation, we employed an in vitro assay in which conjugation of YFP-SUMO to CFP-RanGAPtail can be measured as a change in FRET efficiency if the E1 and E2 enzymes as well as ATP are provided (Bossis et al., 2005). Addition of micromolar concentrations of PPi caused a strong inhibition of CFP-RanGAPtail SUMOylation by the human E1 (Uba2/Aos1) and E2 (Ubc9) which could be released by addition of a soluble pyrophosphatase (Figure 6B + 6C). To determine the mode of inhibition, we determined Vmax and Km in the absence as well as in the presence of PPi (Figure 6B) leading to the conclusion that inhibition of E1E2 activity by PPi follows a mixed mode (Figure 6D, Figure 6—figure supplement 1). We purified the Arabidopsis E1 heterodimer SAE1b SAE2 but could not detect activity in the FRET assay (Figure 6—figure supplement 2) and thus analysed SAE2 ~SUMO thioester formation in the presence and absence of PPi. In accordance with the results for the human enzyme, the Arabidopsis SUMO E1-activity is inhibited by 10 µM PPi (Figure 6E).
Figure 6 with 2 supplements see all
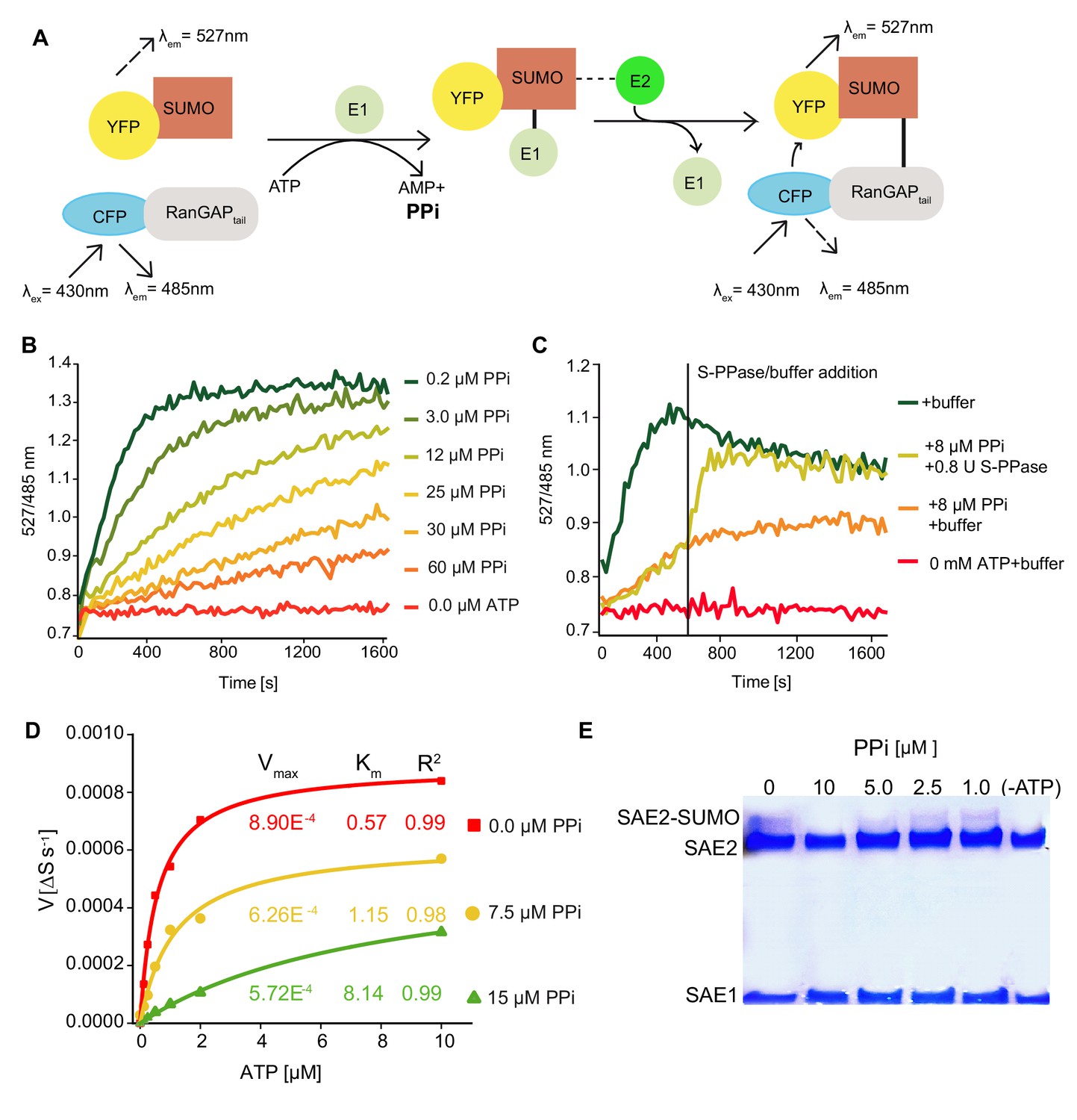
PPi regulates SUMOylation activity in vitro.
(A) Schematic illustration of the FRET-based sumoylation assays. Upon addition of ATP, the human SUMO E1 activating enzyme Aos1/Uba2 and the E2 conjugating enzyme Ubc9 form an isopeptide bond between the CFP-tagged human model substrate GAPtail and YFP-tagged mature SUMO. This can be detected via FRET measurements: Following the excitation of CFP, energy is transferred onto YFP. YFP and CFP emission are recorded upon excitation at 430 nm. Measurements are calculated as the ratio of the λem (YFP-SUMO 527 nm) to λex (CFP-RanGAPtail, 485 nm). (B) PPi titration showing that the increasing PPi concentration inhibits the SUMOylation activity. 1 mM ATP used for all the measurements. (C) In vitro SUMOylation assay showing that the E. coli soluble PPase is able to remove the inhibitory effect of PPi. After 10 min of measurement, one of the 8 µM PPi containing wells were supplied with 0.8 U of E.coli soluble PPase and control buffer was added to the rest of the wells. Measurements were continued for 20 more minutes. (D) PPi addition results in mixed inhibition of SUMOylation activity. Michaelis-Menten fittings of the measurements shown in Figure 6—figure supplement 1. Fittings are done in Origin software and Vmax and Km are calculated accordingly. (E) In vitro thioester bond formation assay showing Arabidopsis E1 (SAE2/SAE1a) and SUMO conjugation under different PPi concentrations. (B–D) Experiments were repeated four times, one representative experiment is shown.
-
Figure 6—source data 1
(B) Ratio values (527/485 nm) of the FRET based SUMOylation showing the effect of the increasing amount of PPi concentrations.
(C) Ratio values (527/485 nm) of the FRET based SUMOylation assay showing that theE. colisoluble PPase is able to remove the inhibitory effect of PPi. (D) Calculation of the Vmax and Km of SUMOylation activity upon PPi inhibition based on the data shown in Figure 6—figure supplement 1.
- https://doi.org/10.7554/eLife.44213.023
Discussion
It has long been assumed that the combined action of V-ATPase and V-PPase enables plants to maintain transport into the vacuole even under stressful conditions (Maeshima, 2000). We have shown previously that the increased activity of the V-ATPase during cold acclimation is largely dependent on the presence of the V-PPase (Kriegel et al., 2015). During cold acclimation fugu5 mutants thus should not be able to adjust their tonoplast proton-pumping activity to the increased demand caused by the accumulation of soluble sugars, organic acids and other osmoprotectants in the vacuole (Schulze et al., 2012). We show here that lack of the V-PPase indeed limits cold acclimation severely. However complementation by overexpression of the soluble pyrophosphatase PPa5 shows clearly that this phenotype is not caused by a reduced proton-gradient limiting cold-induced accumulation of solutes in the vacuole (Figure 1). Accumulation of PPi has been shown to be causative for the developmental phenotype of fugu5 seedlings (Ferjani et al., 2018; Ferjani et al., 2011) and our results show that this also applies to the freezing tolerance and heat stress phenotypes caused by the lack of AVP1 that we report here for the first time. Although overexpression of AVP1 has been shown to result in increased stress tolerance and yield in multiple crop plants, reduced stress tolerance of fugu5 mutants has so far not been reported. The fact that the seedling phenotype observable during the heterotrophic phase of fugu5 seedlings could be rescued by supply of exogenous sucrose pointed to an inhibition of gluconeogenesis. The Glc1P/UDP-Glc reaction is reversible and it has been shown that UGP-Glc pyrophosphorylase is a major target of PPi-inhibition during seedling establishment (Ferjani et al., 2018). Similarly, PPi accumulation could inhibit sugar accumulation during cold acclimation but the fact that the early transcriptional response to cold is dampened in the fugu5 mutant is not easily explained solely by a shift in sugar metabolism (Gutiérrez-Luna et al., 2018). PPi is not only released by many anabolic reactions but also by E1 enzymes that initiate the attachment of ubiquitin or ubiquitin-like proteins (UBLs) including SUMO. Activation of UBLs requires ATP and occurs via carboxy-terminal adenylation and thiol transfer leading to the release of AMP and PPi and would thus be prone to inhibition by PPi accumulation (Desterro et al., 1999; Schulman and Harper, 2009). The MYC-like bHLH transcriptional activator ICE1 is subject to ubiquitination-mediated proteolysis under ambient temperature that is counteracted by SUMOylation during the cold response (Miura and Hasegawa, 2008). We have shown here that the compromised cold acclimation of fugu5 is likely caused by the failure to stabilize ICE1 and that the overall levels of SUMO-conjugates that rapidly increase upon cold exposure in the wild-type fail to increase in fugu5 (Figure 3). It remains to be determined if accumulation of PPi has a similar effect on ubiquitination. However, reduced ubiquitination would stabilize ICE1 and would result in increased cold tolerance. As we cannot exclude that the altered sugar metabolism of fugu5 directly contributes to freezing tolerance or indirectly impinges SUMOylation during cold acclimation, we extended our analysis to the heat stress response. The rapid and reversible accumulation of SUMO conjugates is one of the fastest molecular responses observed during heat stress (Kurepa et al., 2003; Rytz et al., 2018). The fact that this response is dampened in both plants and yeast when PPi accumulates (Figures 4 and 5) argues strongly against a secondary metabolic effect. Evidence for a direct inhibitory effect of PPi on SUMOylation was obtained in an in vitro FRET-based assay that allowed us to determine that the SUMOylation of RanGAP catalysed by human E1 and E2 enzymes was inhibited by micromolar concentrations of PPi following a mixed mode of inhibition (Figure 6). Although we cannot exclude that PPi could inhibit the action of the E2 enzyme, the reaction catalysed by the heterodimeric E1 activating enzyme releases PPi and is thus most likely inhibited when PPi accumulates. Indeed, we could show that E1 subunit SAE2 ~SUMO thioester formation is inhibited in the presence of micromolar PPi, raising the question how exactly PPi inhibits E1-activity (Figure 6).
For adenylation of the SUMO C-terminus to occur, the E1 enzyme adopts an open conformation that allows binding of ATP. In this conformation, the catalytic cysteine of E1 is too far away and unavailable to become linked to SUMO. Thioester bond formation between E1 and SUMO requires structural remodelling to a closed conformation in which the catalytic cysteine moves adjacent to the C terminus of SUMO ~AMP, via unfolding of structures associated with ATP binding and SUMO adenylation (Lois and Lima, 2005; Olsen et al., 2010). It has been suggested that active site remodelling pushes the E1 reaction forward by promoting the release of pyrophosphate to prevent the reverse reaction, the attack of the adenylate by pyrophosphate leading to the reformation of ATP. Not only is the adenylation step rate limiting, once the thioester bond is formed and AMP is released, E1 switches back to the open conformation and a second adenylation reaction occurs, resulting in the formation of a ternary complex, with an E1 molecule binding to one SUMO molecule at the adenylation active site and to a second via a thioester bond through the catalytic cysteine (Olsen et al., 2010). As E1 enzymes are potential targets for therapeutic intervention in cancer and other diseases understanding their enzymatic activity as well as inhibitory mechanisms at the atomic level may provide leads for the development of novel drugs. A novel allosteric inhibitor that targets a cryptic pocket distinct from the active site and locks the enzyme in a previously unobserved inactive conformation has recently been identified (Lv et al., 2018) and it will be of great interest to determine how accumulation off PPi affects the conformation of E1.
Although the exact mechanism remains to be determined, the fact that E1 activity is classically measured as ATP:PPi (Haas et al., 1982; Haas and Rose, 1982) exchange clearly reflects that inhibition of E1 enzymes by PPi is not novel per se. Although cytosolic PPi concentrations reported in the literature, in particular for plants, strongly suggest that relevant concentrations occur not only in mutant backgrounds or under stress conditions the relevance of inhibition by PPi in vivo has so far not been addressed. Cytosolic PPi concentrations of 0.2–0.3 mM as reported for spinach leaves (Weiner et al., 1987) would clearly not be compatible with E1 activity suggesting that PPi levels are maintained at substantially lower levels at least in the immediate environment of E1 enzymes. Many nuclear proteins are modified by SUMOylation (Rytz et al., 2018) and the SUMO conjugation complex self-assembles into nuclear bodies (Mazur et al., 2019). Information regarding the nuclear concentration of PPi is lacking, but the fact that DNA and RNA synthesis occurs against such high concentrations of PPi argues not only that soluble pyrophosphatases play an important role in the nucleus but could also suggest that nuclear PPase activity is higher than in the cytosol. However, the fact that a quadruple knockout mutant lacking four of five PPa-isoforms showed no obvious growth phenotype whereas the combined loss of the H+-PPase AVP1 and a single PPa-isoform causes severe dwarfism due to high PPi concentrations (Segami et al., 2018) shows clearly that cytosolic and nuclear pools of PPi are controlled by the combined action of s PPase and H+-PPase.
At least for plants, converting the substantial energy present in PPi into a proton-gradient seems preferable to releasing it as heat and the soluble PPases might thus only function as emergency valves. But is accumulation of PPi to inhibitory levels only occurring in mutant backgrounds or is there evidence that it is actively prevented under stress conditions in the wild-type? The fact that four PPa-isoforms are transcriptionally up-regulated during the first six hours of the cold acclimation response (Figure 2) indicates that control of PPi levels is an integral part of the cold stress response and it remains to be determined if this is also true for other responses in particular for heat stress. Constitutive overexpression of AVP1 has been shown to cause increased growth of diverse crop plants under various abiotic stress conditions. Greater vacuolar ion sequestration, increased auxin transport, enhanced heterotrophic growth, and increased source to sink transport of sucrose have all been proposed to explain individual aspects of the phenotypes observed in plants lacking or over-expressing AVP1 (Park et al., 2005; Pasapula et al., 2011; Schilling et al., 2017; Yang et al., 2014). Here, we propose modulation of SUMOylation by cellular pyrophosphate levels as a unifying hypothesis that might explain both, the stress-related as well as the developmental aspects of the multifaceted AVP1 loss- and gain-of function phenotypes. Although our hypothesis needs further experimental validation in particular regarding the developmental phenotypes, it seems obvious that a combination of tissue-specific and inducible expression of PPi-hydrolysing enzymes might turn out to be an efficient way of generating stress-tolerant crops for the future.
Materials and methods
Key resources table
| Reagent type (species) or resource | Designation | Source or reference | Identifiers | Additional information |
|---|---|---|---|---|
| Gene (Arabidopsis thaliana) | AtAVP1 | TAIR: AT1G15690 | ||
| Gene (Arabidopsis thaliana) | AtPPa1 | TAIR: AT1G01050 | ||
| Gene (Arabidopsis thaliana) | AtPPa2 | TAIR: AT2G18230 | ||
| Gene (Arabidopsis thaliana) | AtPPa4 | TAIR: AT3G53620 | ||
| Gene (Arabidopsis thaliana) | AtPPa5 | TAIR: AT4G01480 | ||
| Gene (Arabidopsis thaliana) | AtSUMO1 | TAIR: AT4G26840 | ||
| Gene (Arabidopsis thaliana) | AtICE1 | TAIR: AT3G26744 | ||
| Gene (Arabidopsis thaliana) | AtSAE1 | TAIR: AT4G24940 | ||
| Gene (Arabidopsis thaliana) | AtSAE2 | TAIR: AT2G21470 | ||
| Gene (Saccharomices cerevisiae) | ScIPP1 | SGD: YBR011C | ||
| Strain, strain background (Agrobacterium tumefaciens) | ASE | Lampropoulos et al., 2013 | pSOUP+ | |
| Strain, strain background (Saccharomices cerevisiae) | W303 | Szoradi et al., 2018 | SSY122 | |
| Strain, strain background (Saccharomices cerevisiae) | IPP1prΔ::HIS3- GAL1pr-HA-IPP1 | this paper | SSY2542 | Sebastian Schuck lab |
| Genetic reagent (Arabidopsis thaliana) | fugu5-1 | Ferjani et al., 2011 | ||
| Genetic reagent (Arabidopsis thaliana) | fugu5-3 | Ferjani et al., 2011 | ||
| Genetic reagent (Arabidopsis thaliana) | ice1-2 | Nottingham Arabidopsis Stock Centre (NASC) | SALK_003155 | |
| Genetic reagent (Arabidopsis thaliana) | ppa1 | Segami et al., 2018; NASC | SAIL_251_D07 | |
| Genetic reagent (Arabidopsis thaliana) | ppa1,4 | this paper | SAIL_251_D07, SAIL_916_C08 | Cross between the mutants ppa1 and ppa4 described in Segami et al., 2018 Masayoshi Maeshima lab |
| Genetic reagent (Arabidopsis thaliana) | ppa1,2,4,5 | Segami et al., 2018 | SAIL_251_D07, SAIL_618_H05, SAIL_916_C08, SALK_014647 | |
| Biological sample (Arabidopsis thaliana) | AVP1:IPP1/fugu5-1 | Ferjani et al., 2011 | AVP1 promoter:IPP1 coding sequence; fugu5-1 mutant background | |
| Biological sample (Arabidopsis thaliana) | UBQ:AVP1 #18–4 | Kriegel et al., 2015 | UBQ promoter:AVP1 coding sequence; Col-0 wild-type background | |
| Biological sample (Arabidopsis thaliana) | UBQ:PPa5-GFP/Col-0 | this paper, | UBQ promoter:PPa5-GFP coding sequence; Col-0 wild-type background. Karin Schumacher lab | |
| Biological sample (Arabidopsis thaliana) | UBQ:PPa5-GFP/fugu5-1 | this paper, | UBQ promoter:PPa5-GFP coding sequence; fugu5-1 wild-type background. Karin Schumacher lab | |
| Antibody | Anti-SUMO1 (rabbit polyclonal) | Agrisera | AS08308 | (1:1000) |
| Antibody | Anti-ICE1 (rabbit polyclonal) | Agrisera | AS163971 | (1:1000) |
| Antibody | Anti-cFBPase (rabbit polyclonal) | Agrisera | AS04043 | (1:5000) |
| Antibody | Anti-rabbit-HRP (Goat polyclonal) | Promega | W401B | (1:10000) |
| Antibody | Anti-IPP1 (rabbit polyclonal) | Antibodies- online GmbH | ABIN459215 | (1:1000) |
| Antibody | Anti-PGK1 (mouse monoclonal) | Abcam | AB113687 | (1:100000) |
| Antibody | Anti-mouse-HRP (sheep polyclonal) | GE Healthcare UK | NXA931 | (1:5000) |
| Antibody | Anti-V-PPase (rabbit polyclonal) | Agrisera | AS121849 | (1:10000) |
| Antibody | Anti-VHA-C (rabbit polyclonal) | Karin Schumacher lab | Schumacher et al., 1999 | (1:2000) |
| Antibody | Anti-GFP (rabbit polyclonal) | Karin Schumacher lab | Roth et al., 2018 | (1:10000) |
| Recombinant DNA reagent | UBQ:PPa5-GFP | this paper, | vector-promoter:tagged-protein construct. Karin Schumacher lab | |
| Recombinant DNA reagent | pET28b(+)−6xHIS -AtSUMO1(1–93) | this paper, | vector-tagged-protein construct. Karin Schumacher lab | |
| Recombinant DNA reagent | pET28-6xHIS-AtSAE1 | this paper, | vector-tagged-protein construct. Frauke Melchior lab | |
| Recombinant DNA reagent | pET11d-AtSAE2 | this paper | vector-protein construct. Frauke Melchior lab | |
| Recombinant DNA reagent | pFA6a-His3M × 6- PGAL1 | this paper, | vector-promoter:tagged-protein construct. Sebastian Schuck lab | |
| Peptide, recombinant protein | CFP-RanGAPtail | Bossis et al., 2005; Werner et al., 2009 | ||
| Peptide, recombinant protein | YFP-SUMO | Bossis et al., 2005; Werner et al., 2009 | ||
| Peptide, recombinant protein | Uba2/Aos1 | Bossis et al., 2005; Werner et al., 2009 | ||
| Peptide, recombinant protein | Ubc9 | Bossis et al., 2005; Werner et al., 2009 | ||
| Peptide, recombinant protein | 6xHis-AtSUMO1 | this paper | For in vitro thioester bond formation assays | |
| Peptide, recombinant protein | AtSAE1 | this paper | For in vitro thioester bond formation assays | |
| Peptide, recombinant protein | AtSAE2 | this paper | For in vitro thioester bond formation assays | |
| Commercial assay or kit | SPL kit | NH DyeAgnostics | Western blot protein quantification kit |
Plant material and growth conditions
Request a detailed protocolArabidopsis thaliana Col-0 ecotype was used in all experiments as control. Description of all mutant and transgenic lines used in this work can be found in the key resources table. Seeds were surface sterilized with ethanol and stratified for 48 hr at 4°C. Seedlings for propagation, microscopy and heat shock tolerance assays were grown on plates with standard growth medium (0.5% Murashige and Skoog (MS), 0.5% phyto agar, and 10 mM MES, pH 5.8) for 10 days under long day conditions (16 hr light/8 hr dark) at 22°C at 125 µmol·m−2·s−1. For heat shock tolerance assays at day 10, treatment plates were exposed to 40°C for 4 hr while the control plates were kept in growth conditions. For freezing tolerance assays, electrolyte leakage assays, PPi and sugar determination,real time RT-PCR, enzyme assays, cell sap measurements and phenotypic assays plants were grown for 6 weeks on soil under short day conditions (10 hr light/14 hr dark) at 22°C at 125 µmol·m−2·s−1. In the cases where cold acclimation was required plants were incubated for 4 days at 4°C while the untreated plants were maintained in the same conditions as the growth period. To test the ICE1 antibody, Col-0, fugu5-1 and ice1-2 were grown on soil under long day conditions for 3 weeks at 22°C at 125 µmol·m−2·s−1. To determine SUMO and ICE1 protein amounts upon cold and heat treatments, seedlings were grown in liquid culture (0.5% Murashige and Skoog (MS), 0.5% sucrose, 10 mM MES, pH5.8) under long day conditions at 22°C at 125 µmol·m−2·s−1. Growth period was 10 days in 50 ml liquid culture in a 300 ml flask on a horizontal shaker with 100 rpm speed. After 10 days, part of flasks was either subjected to 30 min 40°C or 3 hr 4°C. Control samples were kept at normal growth conditions. For analysis of cotyledon phenotypes seeds were planted in single pots individually after surface sterilization (n = 10). Pictures were taken at day 5.
Construct design and plant transformation
Request a detailed protocolUBQ:PPa5-GFP construct was generated using GreenGate (GG) cloning (Lampropoulos et al., 2013). The 1097 base pairs coding sequence of PPa5 was amplified from Arabidopsis thaliana Col-0 cDNA with primers listed in Supplementary file 1A, attaching BsaI recognition sites and specific GG-overhangs. To prevent cutting, the internal BsaI site was mutated by site directed mutagenesis. Thereafter, the PCR product and the empty entry module (pGGC000) were digested with BsaI, the digestion was purified and then ligated. After test digestion positive clones were checked by sequencing. The final construct was assembled in a GG reaction from modules listed in Supplementary file 1B and transformed into Agrobacterium tumefaciens ASE strain harbouring the pSOUP plasmid. Arabidopsis thaliana ecotype Col-0 and fugu5-1 plants were used for transformation via floral dipping (Clough and Bent, 1998).
Yeast strain generation and growth
Request a detailed protocolTo replace the endogenous IPP1 promoter with the inducible GAL1 promoter and simultaneously introduce an N-terminal HA-tag, plasmid pFA6a-His3M × 6-PGAL1 was amplified with primers Ipp1-F4/Ipp1-R3 (Longtine et al., 1998). The resulting PCR product was used for transformation of a wild type S. cerevisiae W303 strain (SSY122; (Szoradi et al., 2018). Correct promoter replacement in the resulting IPP1prΔ::HIS3-GAL1pr-HA-IPP1 strain (SSY2542) was confirmed by colony PCR and lack of growth on glucose-containing medium. Cells were grown on synthetic complete medium (CSM –Uracil (MP #4511212), Difco yeast nitrogen base (BD #233520), Uracil (Sigma #U1128), Adenine Hemisulfate (Sigma #A9126)) supplemented with appropriate carbon sources. All determinations were done on exponentially growing cells (A600 ≤0.5). To maintain cultures for several hours below an A600 of 0.5, they were diluted with fresh medium every two hours until the end of the experiment (semi-continuous culture). A pre-culture containing galactose was grown at 28°C shaking until A600 0.5, then divided to four, for temperature and carbon resource manipulation: Glucose/28°C, Galactose/28°C, Galactose/40°C, Glucose/40°C. Samples were taken at indicated time points (0, 6, 15 hr). For heat treatment samples were taken from 28°C one hour before the indicated time point and incubated at 40°C for an hour.
Protein preparation and immunoblotting analysis
Request a detailed protocolTo determine total SUMOylation amount in Arabidopsis, total proteins were extracted from liquid grown wt, V-PPase mutants, UBQ:PPa5/fugu5-1 and UBQ:AVP1 as described in (Castaño-Miquel et al., 2013). 15 µg protein was loaded to 7.5%, 1.5 mM SDS-gels. Anti-SUMO1 (1:1000; Agrisera) was used as primary antibody. To measure ICE1 protein, Col-0 and fugu5-1 and ice1-2 were used for total protein extraction as described in Castaño-Miquel et al. (2013). Anti-ICE1 (1:1000; Agrisera) was used as primary antibody. Anti-cFBPase (Agrisera; 1:5000) was used as loading control for both SUMO and ICE1. For all immunoblots, HRP-anti-rabbit was used as secondary antibody (1:10000; Promega). To determine the amount of soluble pyrophosphatase and the amount of total SUMOylated proteins in yeast, total protein extraction was performed as described in Szoradi et al. (2018) with addition of 20 mM NEM. 15 µg protein was loaded to 7.5%, 1.5 mM SDS-gels. Anti-IPP1 (Antibodies-online GmbH, 1:1000) and anti-SUMO1 (1:1000; Agrisera) were used as primary antibodies. HRP anti-rabbit was used as secondary antibody. Anti-PGK1 (Abcam, 1:100000) was used as the loading control. An anti-mouse antibody was used as secondary antibody (GE Healtcare UK, 1:5000).
To determine protein levels of V-ATPase and V-PPase we prepared tonoplast membrane extracts of Col-0, fugu5-1 and UBQ:AVP1. The primary antibody against the V-PPase was purchased from Agrisera (1:10,000) and the primary antibody against VHA-C is as previously described (Schumacher et al., 1999). To determine the UBQ:PPa5-GFP levels in Col-0 and fugu5-1 background soluble proteins were extracted as described in tonoplast vesicle preparation section. Anti-GFP (1:10000) was used as primary antibody. An internal control from SPL kit was used for normalization (NH DyeAgnostics). To measure ICE1 protein, Col-0 and fugu5-1 were grown in liquid culture. Material was split in two for total protein extraction, same buffer described in Castaño-Miquel et al., 2013 was used for one part, and same buffer without NEM and with addition of 5 mM DTT used for the other. Anti-ICE1 (1:1000; Agrisera) was used as primary antibody. For all immunoblots, HRP-anti-rabbit was used as secondary antibody (1:10000; Promega). Imaging was carried out using a cooled CCD camera system (Intas ADVANCED Fluoreszenz u. ECL Imager). Western blots were quantified with Fiji (based on ImageJ 1.47 t).
Tonoplast vesicle preparation and enzyme assays
Request a detailed protocolRosette leaf material (75 g) from plants grown under short day conditions was harvested. The leaf material was homogenized in homogenization buffer containing 0.4 M mannitol, 0.1 M Tris, 10% (vol/vol) glycerol, 3 mM Na2EDTA, 0.5% (wt/vol) BSA, 5% (vol/vol) PVP-10, 0.5 mM butylated hydroxytoluene, 0.3 mM dibucaine, 5 mM magnesium sulphate, 1 mM PMSF (phenylmethylsulphonylfluoride), 1.3 mM benzamidine and 25 mM potassium metabisulfite. The homogenate was filtered through two layers of miracloth and centrifuged at 10,000 g for 20 min at 4°C. The supernatant was then centrifuged at 100,000 g for 45 min at 4°C. The microsomal membrane pellet was resuspended in resuspension buffer containing 0.4 M mannitol, 6 mM Tris-MES (pH 8) and 10 (vol/vol) glycerol. Soluble part was kept for measuring the soluble pyrophosphatase activity and quantification of PPa5-GFP protein levels. Tonoplast vesicles were obtained by performing a sucrose gradient with 22% sucrose. Centrifugation was performed at 97,000 g for 2 hr. Protein concentrations were determined as reported previously (Bradford, 1976). ATP and PPi hydrolysis was measured at 28°C as described previously (Krebs et al., 2010). Same method was also used for measuring soluble pyrophosphatase activity with soluble proteins. The ATP and PPi-dependent proton transport activities were estimated from the initial rate of ATP-dependent fluorescence quenching in the presence of 3 mM ATP using the fluorescence dye ACMA (9-Amino-6-Chloro-2-Methoxyacridine) with 50 µg enriched tonoplast protein. Excitation wavelength was 415 nm, and emission was measured at 485 nm in Jasco fluorescence spectrometer. V-PPase H+ transport medium includes 25 mM HEPES-BTP (pH 7.2), 250 mM Sorbitol, 1.5 mM MgSO4, 50 mM KCl and 0.3 mM PPi-BTP (pH 7.5) final concentration in 1 ml volume. V-ATPase H+ transport medium includes 10 mM ATP-MES (pH 8.0),0.25 M Mannitol, 3 mM MgSO4, 100 mM TMA-Cl and 1.5 mM ATP-BTP (pH 7.5) final concentration in 1 ml volume.
Cell sap pH measurements
Request a detailed protocolCell sap pH measurements were conducted as previously described (Krebs et al., 2010)
Confocal microscopy
Request a detailed protocolLocalization of UBQ:PPa5-GFP construct was determined using a Leica TCS SP5II microscope equipped with a Leica HCX PL APO lambda blue 63.0 3 1.20 UV water immersion objective. GFP was excited at 488 nm using a VIS-argon laser. Fluorescence emission of GFP was detected between 500 and 555 nm. The Leica Application Suite Advanced Fluorescence software was used for image acquisition. Post processing of images were performed using Fiji.
Determination of PPi and soluble sugar levels via ion-chromatography
Request a detailed protocol6 weeks old short day grown rosettes were ground in liquid nitrogen and aliquots of ~200–400 mg were used to quantify PPi and soluble sugars. Compounds were extracted with 0.5 ml ultra-pure water for 20 min at 95°C with vigorous shaking, and insoluble material was removed by centrifugation at 20,800 g for 20 min. PPi was measured using an IonPac AS11-HC (2 mm, Thermo Scientific) column connected to an ICS-5000 system (Thermo Scientific) and quantified by conductivity detection after cation suppression (ASRS-300 2 mm, suppressor current 29–78 mA). Prior separation, the column was heated to 30°C and equilibrated with five column volumes of ultra-pure water at a flow rate of 0.3 ml/min. Soluble sugars were separated on a CarboPac PA1 column (Thermo Scientific) connected to the ICS-5000 system and quantified by pulsed amperometric detection (HPAEC-PAD). Column temperature was kept constant at 25°C and equilibrated with five column volumes of ultra-pure water at a flow rate of 1 ml min-1. Data acquisition and quantification was performed with Chromeleon 7 (Thermo Scientific).
Freezing tolerance assay
Request a detailed protocolPlant freezing tolerance was determined with 6 weeks old short-day grown plants. For cold acclimation, 6-week-old plants were incubated at 4°C for 4 days with same photoperiod. Non-acclimated plants were kept at 22°C during this period. Plants were wetted thoroughly to promote freezing, then placed in a controlled temperature chamber (Polyklima, MN2-WLED). First they were kept at 0°C for 1 hr. Afterwards, they were subjected to temperatures from −1 to −10°C, reduced 1°C every 30 min. After thawing at 4°C overnight, plants were moved back to 22°C. Images were taken before cold treatment and 1 week after the freezing treatment. Dead and alive leaves were counted after the photos were taken.
Electrolyte leakage from leaves
Request a detailed protocolElectrolyte leakage was measured from fully developed rosette leaves of 6-week-old plants. For each temperature five leaves were collected from each genotype. Each leaf (5th or 6th rosette leaf) was placed into a tube containing 3 mL deionized water, then placed to 0°C at a temperature-controlled climate chamber. Temperature was decreased by 2°C every hour. At −2°C an ice chip was added to initiate nucleation. Tubes were collected at −2,–4, −6,–8 and −10°C and placed to 4°C to thaw overnight. Next day 2 ml deionized water was added and tubes were placed overnight on a horizontal shaker (100 rpm) at 4°C. Conductivity after freezing was measured with a conductivity meter (Mettler-Toledo, FiveEasy), which was calibrated with the Mettler-Toledo Buffer solution 1413 µS. Then, samples were placed to a 100°C water bath and boiled for 2 hr. Conductivity was again measured after boiling. Ion leakage was determined as the percent ratio of the measurement of conductivity before and after boiling.
RNA isolation and cDNA synthesis
Request a detailed protocolFor the analysis of transcript levels 6 weeks old Col-0, fugu5-1 and UBQ:PPa5-GFP/fugu5-1 was collected after exposure to 4°C for indicated time points. RNA was isolated using the RNeasy Plant Mini Kit (Qiagen) according to manufacturer's instructions. cDNA was synthesized from 1 μg of total RNA using M-MuLV reverse transcriptase (Thermo) and an oligo dT primer.
Real-time RT PCR
Request a detailed protocolFor quantitative analysis of gene expression real-time RT PCR was applied. cDNA samples were diluted 1:50 in nuclease-free water. Real-time PCR reactions were performed using the DNA Engine Opticon System (DNA Engine cycler and Chromo4 detector, BioRad) and SG qPCR mastermix 2X (Roboklon). The real-time PCR reaction mixture with a final volume of 20 μl contained 0.5 μM of each forward and reverse primer, 10 μl SYBR Green Mix, 4 μl cDNA and 4 μl of RNase-free water. The thermal cycling conditions were composed of an initial denaturation step at 95°C for 15 min followed by 40 cycles at 95°C for 15 s, 60°C for 30 s and 72°C for 15 s and ended with a melting curve. For the analysis of each sample three analytical replicas were used. Target genes were normalized to the expression of Actin2. Primer sequences are listed in Supplementary file 1C.
In vitro FRET- based SUMOylation assay
Request a detailed protocolSUMOylation of CFP-RanGAPtail with YFP-SUMO was carried out using a FRET-based high-throughput assay as previously described with minor changes (Werner et al., 2009; Bossis et al., 2005). Final concentrations of the FRET components were Uba2/Aos1 (E1, 20 nM), Ubc9 (E2, 30 nM), YFP-SUMO and CFP-RanGAPtail (300 nM). ATP substrate was prepared as a stock solution of 300 mM ATP-BTP (pH 8.0). 1 mM ATP was used for the assays unless stated otherwise. For the PPi application, 30 mM PPi-BTP (pH 7.5) stock solution was prepared. To determine the effects of PPi hydrolysis on the FRET assay, 0.8U E. coli inorganic pyrophosphatase (NEB) was used and the buffer solution that the pyrophophatase includes was added to the control wells (20 mM Tris-HCl, 100 mM NaCl, 1 mM Dithiothreitol, 0.1 mM EDTA, 50% Glycerol, pH 8.0). Michealis-Menten fittings and Vmax and Km calculations were done in Origin software according to the ATP titration (0–10 µM) performed with different PPi concentrations (0, 7.5 and 15 µM).
Cloning, expression and protein purification of Arabidopsis E1 ligase and SUMO1
Request a detailed protocolConjugation-competent AtSUMO1 (1–93) was amplified from cDNA, using the primers AtSUMO1-NdeI-Fw and AtSUMO1-XhoI-Rv, and cloned in the bacteria expression vector pET28b(+). The coding sequence of SAE1a was amplified from A. thaliana cDNA ligated into pET28a via NheI and BamHI sites in-frame behind the coding sequence for a 6xHis-tag. SAE2 was amplified from A. thaliana cDNA and ligated into the pCR-Blunt II-TOPO vector. pET11d was cut with BamHI and the TOPO-vector was cut with NheI. Both linear DNA fragments were blunt ended with T4 polymerase. Both DNA fragments were subsequently restricted with NcoI. The DNA fragment carrying the SAE2 coding sequence was ligated into pET11d via the NcoI cohesive end and the blunt end. Recombinant proteins were purified as previously described (Werner et al., 2009).
In vitro E1-Thioester assay
Request a detailed protocolE1-Thioester assay was performed as previously described in Castaño-Miquel et al. (2013) with addition of final concentrations of 1–10 µM PPi. 1 mM of ATP was used for all reactions unless stated otherwise.
No index entries found.
Data availability
All data generated or analysed during this study are included in the manuscript and supporting files. Source data files have been provided.
References
-
BookA Fluorescence Resonance Energy Transfer‐Based Assay to Study SUMO Modification in SolutionMethods in EnzymologyAcademic Press.
-
SUMO, a heavyweight player in plant abiotic stress responsesCellular and Molecular Life Sciences 69:3269–3283.https://doi.org/10.1007/s00018-012-1094-2
-
Cold stress regulation of gene expression in plantsTrends in Plant Science 12:444–451.https://doi.org/10.1016/j.tplants.2007.07.002
-
Transformation of arabidopsis thalianaThe Plant Journal: For Cell and Molecular Biology 16:735–743.https://doi.org/10.1046/j.1365-313x.1998.00343.x
-
Identification of the enzyme required for activation of the small ubiquitin-like protein SUMO-1Journal of Biological Chemistry 274:10618–10624.https://doi.org/10.1074/jbc.274.15.10618
-
BookRegulation of PPi Levels Through the Vacuolar Membrane H+-PyrophosphataseIn: Lüttge U, Beyschlag W, Cushman J, editors. Progress in Botany, 75. Berlin, Heidelberg: Springer Berlin Heidelberg. pp. 145–165.https://doi.org/10.1007/978-3-642-38797-5_5
-
Sumoylation: a regulatory protein modification in health and diseaseAnnual Review of Biochemistry 82:357–385.https://doi.org/10.1146/annurev-biochem-061909-093311
-
Ubiquitin-activating enzyme. mechanism and role in protein-ubiquitin conjugationThe Journal of Biological Chemistry 257:2543–2548.
-
The mechanism of ubiquitin activating enzyme. A kinetic and equilibrium analysisThe Journal of Biological Chemistry 257:10329–10337.
-
Defining the SUMO-modified proteome by multiple approaches in saccharomyces cerevisiaeJournal of Biological Chemistry 280:4102–4110.https://doi.org/10.1074/jbc.M413209200
-
BookBiological Production of PPiIn: Heinonen J. K, editors. Biological Role of Inorganic Pyrophosphate. Boston: Springer US. pp. 1–28.https://doi.org/10.1007/978-1-4615-1433-6_1
-
Protein modification by SUMOAnnual Review of Biochemistry 73:355–382.https://doi.org/10.1146/annurev.biochem.73.011303.074118
-
On the metabolic significance of phosphorolytic and pyrophosphorolytic reactionsHorizons in Biochemistry and Biophysics pp. 251–264.
-
The small ubiquitin-like modifier (SUMO) protein modification system in arabidopsis. accumulation of SUMO1 and -2 conjugates is increased by stressThe Journal of Biological Chemistry 278:6862–6872.https://doi.org/10.1074/jbc.M209694200
-
Yeast PPA2 gene encodes a mitochondrial inorganic pyrophosphatase that is essential for mitochondrial functionThe Journal of Biological Chemistry 266:12168–12172.
-
Vacuolar H+-pyrophosphataseBiochimica Et Biophysica Acta (BBA) - Biomembranes 1465:37–51.https://doi.org/10.1016/S0005-2736(00)00130-9
-
Regulation of cold signaling by sumoylation of ICE1Plant Signaling & Behavior 3:52–53.https://doi.org/10.4161/psb.3.1.4865
-
AVP1: one protein, many rolesTrends in Plant Science 22:154–162.https://doi.org/10.1016/j.tplants.2016.11.012
-
Ubiquitin-like protein activation by E1 enzymes: the apex for downstream signalling pathwaysNature Reviews Molecular Cell Biology 10:319–331.https://doi.org/10.1038/nrm2673
-
Inorganic pyrophosphatase defects lead to cell cycle arrest and autophagic cell death through NAD+ depletion in fermenting yeastJournal of Biological Chemistry 288:13082–13092.https://doi.org/10.1074/jbc.M112.439349
-
Expression of E. coli inorganic pyrophosphatase in transgenic plants alters photoassimilate partitioningThe Plant Journal : For Cell and Molecular Biology 2:571–581.
-
PLANT COLD ACCLIMATION: freezing tolerance genes and regulatory mechanismsAnnual Review of Plant Physiology and Plant Molecular Biology 50:571–599.https://doi.org/10.1146/annurev.arplant.50.1.571
-
Subcellular compartmentation of pyrophosphate and alkaline pyrophosphatase in leavesBiochimica Et Biophysica Acta (BBA) - Bioenergetics 893:13–21.https://doi.org/10.1016/0005-2728(87)90143-5
-
Performing in vitro sumoylation reactions using recombinant enzymesMethods in Molecular Biology 497:187–199.https://doi.org/10.1007/978-1-59745-566-4_12
Article and author information
Author details
M Görkem Patir-Nebioglu

Funding
Deutsche Forschungsgemeinschaft (SCHU1151/13-1)
- Karin Schumacher
Deutsche Forschungsgemeinschaft (AN1323/1-1)
- Zaida Andrés
Deutsche Forschungsgemeinschaft (SFB1036 (TP15))
- Frauke Melchior
The funders had no role in study design, data collection and interpretation, or the decision to submit the work for publication.
Acknowledgements
We are grateful to Beate Schöfer, Barbara Jesenofsky and Anne Newrly for excellent technical assistance. This work was supported by DFG grants SCHU1151/13-1 to KS, AN1323/1-1 to ZA and SFB1036 (TP15) to FM. SS acknowledges support through DFG grant EXC 81.
Version history
- Received: December 7, 2018
- Accepted: February 5, 2019
- Version of Record published: February 20, 2019 (version 1)
Copyright
© 2019, Patir-Nebioglu et al.
This article is distributed under the terms of the Creative Commons Attribution License, which permits unrestricted use and redistribution provided that the original author and source are credited.
Metrics
-
- 5,372
- views
-
- 410
- downloads
-
- 1
- citations
Views, downloads and citations are aggregated across all versions of this paper published by eLife.
Download links
A two-part list of links to download the article, or parts of the article, in various formats.
Downloads (link to download the article as PDF)
Open citations (links to open the citations from this article in various online reference manager services)
Cite this article (links to download the citations from this article in formats compatible with various reference manager tools)
RETRACTED: Pyrophosphate modulates plant stress responses via SUMOylation
eLife 8:e44213.
https://doi.org/10.7554/eLife.44213
Further reading
-
- Biochemistry and Chemical Biology
- Neuroscience
In most mammals, conspecific chemosensory communication relies on semiochemical release within complex bodily secretions and subsequent stimulus detection by the vomeronasal organ (VNO). Urine, a rich source of ethologically relevant chemosignals, conveys detailed information about sex, social hierarchy, health, and reproductive state, which becomes accessible to a conspecific via vomeronasal sampling. So far, however, numerous aspects of social chemosignaling along the vomeronasal pathway remain unclear. Moreover, since virtually all research on vomeronasal physiology is based on secretions derived from inbred laboratory mice, it remains uncertain whether such stimuli provide a true representation of potentially more relevant cues found in the wild. Here, we combine a robust low-noise VNO activity assay with comparative molecular profiling of sex- and strain-specific mouse urine samples from two inbred laboratory strains as well as from wild mice. With comprehensive molecular portraits of these secretions, VNO activity analysis now enables us to (i) assess whether and, if so, how much sex/strain-selective ‘raw’ chemical information in urine is accessible via vomeronasal sampling; (ii) identify which chemicals exhibit sufficient discriminatory power to signal an animal’s sex, strain, or both; (iii) determine the extent to which wild mouse secretions are unique; and (iv) analyze whether vomeronasal response profiles differ between strains. We report both sex- and, in particular, strain-selective VNO representations of chemical information. Within the urinary ‘secretome’, both volatile compounds and proteins exhibit sufficient discriminative power to provide sex- and strain-specific molecular fingerprints. While total protein amount is substantially enriched in male urine, females secrete a larger variety at overall comparatively low concentrations. Surprisingly, the molecular spectrum of wild mouse urine does not dramatically exceed that of inbred strains. Finally, vomeronasal response profiles differ between C57BL/6 and BALB/c animals, with particularly disparate representations of female semiochemicals.




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}