Multilocus Sequence Typing of Non-JP2 Serotype b Aggregatibacter actinomycetemcomitans Strains of Ghanaian and Swedish Origin
 Rolf Claesson1
Rolf Claesson1  Anders Johansson
Anders Johansson Anders Esberg
Anders Esberg Dorte Haubek
Dorte Haubek Jan Oscarsson
Jan Oscarsson- 1Division of Oral Microbiology, Department of Odontology, Umeå University, Umeå, Sweden
- 2Division of Molecular Periodontology, Department of Odontology, Umeå University, Umeå, Sweden
- 3Department of Odontology, Umeå University, Umeå, Sweden
- 4Section for Paediatric Dentistry, Department of Dentistry and Oral Health, Aarhus University, Aarhus, Denmark
Objective and Methods: The Gram-negative bacterium, Aggregatibacter actinomycetemcomitans is associated with periodontitis affecting young individuals. The geographic dissemination of the highly leukotoxic JP2 genotype of serotype b of this species was previously studied by multilocus sequence typing (MLST). Here, we have used MLST to genetically characterize non-JP2 genotype strains of serotype b, isolated from individuals living in Ghana (n=41), and in Sweden (n=13), respectively.
Results: The MLST analysis revealed a total of nine sequence types (ST). Both Ghanaian and Swedish isolates were distributed in ST 1-3. ST 5 and 6 were only identified among the Ghanaian strains, whereas ST 4, 7, 8 and 9 were uniquely represented among the Swedish strains. Previously, we characterized these non-JP2 genotype strains of A. actinomycetemcomitans serotype b by arbitrarily-primed (AP)-PCR, which distributed them into three groups, AP-PCR type 1, 2, and 3, respectively. AP-PCR type 1 strains are generally highly leukotoxic, and are associated with progression of periodontal attachment loss. As AP-PCR type 1 includes both JP2 genotype strains and a proportion of non-JP2 genotype strains of serotype b, a straightforward diagnostic procedure has been sought. This has revealed a gene, cagE, which appears to be conserved only in this AP-PCR type. According to our results, MLST was not a highly discriminatory method to identify AP-PCR type 1, as strains of this AP-PCR type could be found within three different ST: ST 2, ST 3 and ST 8.
Conclusion: According to MLST, a geographic dissemination of non-JP2 genotype A. actinomycetemcomitans serotype b appears to exist. However, aiming to identify carriers of AP-PCR type 1, non-JP2 genotype serotype b, PCR with cagE-specific primers is likely the most efficient diagnostic procedure known today.
Introduction
Aggregatibacter actinomycetemcomitans produces a toxin, which associates this bacterium with periodontitis affecting young individuals (Haubek et al., 2008; Henderson et al., 2010; Könönen and Muller, 2014; Fine et al., 2020). Since the toxin causes lethal effects on leukocytes of different types, it is considered a leukotoxin (LtxA). The production and release of LtxA is affected by alterations in the promoter region of the ltxCABD operon (Kolodrubetz et al., 1989; Spitznagel et al., 1991). Of the seven serotypes of A. actinomycetemcomitans (a-g), a specific variant of serotype b has a deletion of 530 base pairs (bp) in the promoter region (Brogan et al., 1994). This variant is genetically different from those having a full-length promoter, and is since long referred to as the JP2 genotype (Haubek, 2010). As a consequence, all variants of A. actinomycetemcomitans with a full-length leukotoxin gene promoter are typically referred to as non-JP2 genotype. In serotype b strains, also additional ltxCABD promoter types have been identified, with either deletions of alternative regions of the promoter, or with insertions in it (He et al., 1999; Claesson et al., 2015; Claesson et al., 2020).
The JP2 genotype of A. actinomycetemcomitans serotype b is highly leukotoxic (Tsai et al., 2018), and carriers of this specific type are at substantial risk for rapid loss of tooth-supporting tissues, including alveolar bone (Bueno et al., 1998; Haraszthy et al., 2000; Haubek et al., 2008; Burgess et al., 2017). While the JP2 genotype appears to be distributed only in serotype b of A. actinomycetemcomitans, the non-JP2 genotype is found among all serotypes (a-g). Non-JP2 genotype strains of A. actinomycetemcomitans are generally low-leukotoxic. However, a subgroup of non-JP2 genotype strains of A. actinomycetemcomitans serotype b that exhibit high leukotoxicity, and has enhanced association with periodontal disease progression, has been identified (Asikainen et al., 1995; Höglund Åberg et al., 2014).
In addition to leukotoxin promoter typing, A. actinomycetemcomitans strains have been genotypically characterized by a range of other DNA-based methods (Asikainen and Karched, 2008; Kittichotirat et al., 2016). One of them is arbitrarily-primed (AP)-PCR (van Belkum, 1994), which has revealed that all hitherto tested JP2 genotype strains belong to the same AP-PCR banding pattern group (AP-PCR type 1), whereas for non-JP2 strains a number of different AP-PCR types have been identified (Höglund Åberg et al., 2014; Claesson et al., 2017). Interestingly, a subgroup of non-JP2 genotype strains of A. actinomycetemcomitans serotype b that expresses enhanced levels of LtxA also belongs to AP-PCR type 1, and hence appears to be genetically related to the JP2 genotype of A. actinomycetemcomitans (Höglund Åberg et al., 2014; Johansson et al., 2017). Moreover, all hitherto assessed A. actinomycetemcomitans strains of serotype b belonging to AP-PCR type 1 have the property in common that they carry the gene cagE, which with very few exceptions appears to be present in this AP-PCR type only, and as the gene is absent in the other A. actinomycetemcomitans serotypes, it could be used for diagnostic purposes (Johansson et al., 2017; Johansson et al., 2019).
AP-PCR characterization mirrors genetic variability within bacterial species without requiring the analysis of specific genes, whereas Multilocus Sequence Typing (MLST) is a molecular typing method that is used to characterize bacterial strains in terms of pathogenicity and dissemination (Dogan et al., 1999b; Haubek et al., 2007; Jolley and Maiden, 2014). This is a highly discriminatory typing method based on sequencing of a number of housekeeping genes. MLST of bacterial strains renders so called sequence types (ST), which can be further used in dissemination studies. As judged by the MLST-based characterization, the JP2 genotype of serotype b emerged from the Mediterranean part of Africa about 2400 years ago, and has then subsequently been spread worldwide (Haubek et al., 2007; Rylev and Kilian, 2008).
In the present study, we have carried out MLST-characterization of non-JP2 genotype strains of A. actinomycetemcomitans serotype b from two of our clinical collections, isolated from individuals living in Ghana and Sweden, respectively (Höglund Åberg et al., 2012; Claesson et al., 2017). Thereby, we have been able to compare the genetic variability of non-JP2 genotype strains of A. actinomycetemcomitans serotype b collected from carriers living in two from each other separated continents, to assess if geographic dissemination of such strains may exist. In addition, our aim was to study whether any correlation could be observed between AP-PCR/cagE genotype and MLST–based sequence types of non-JP2 genotype strains of A. actinomycetemcomitans serotype b. To this end, the MLST pattern of each strain was compared with their genetic discrepancies earlier revealed by AP-PCR and cagE genotyping (Höglund Åberg et al., 2014; Claesson et al., 2017; Johansson et al., 2017; Johansson et al., 2019).
Materials and Methods
A. actinomycetemcomitans Strain Collections
A list of all strains used in this work is presented as Supplementary Material (Table S1). A total of 56 non-JP2 genotype strains of A. actinomycetemcomitans serotype b, collected and isolated from subgingival plaque samples, were used in this work. These included 41 collected from adolescents in a Ghanaian (Gh) study cohort (Höglund Åberg et al., 2012), 13 sampled from patients living in Sweden (Sw), and recruited to specialist periodontal treatment (Claesson et al., 2017), and two reference strains named HK908, and HK912. For all strains used in this work, AP-PCR and cagE genotype have earlier been examined, respectively (Höglund Åberg et al., 2014; Claesson et al., 2017; Johansson et al., 2017; Johansson et al., 2019). The 56 strains were grouped into AP-PCR type 1 (n=18, all cagE-positive), AP-PCR type 2 (n=16; all cagE-negative), and AP-PCR type 3 (n=21; 20 cagE-negative). The latter AP-PCR type actually includes AP-PCR types 3 to 11, as defined earlier (Claesson et al., 2017; Johansson et al., 2019). One strain, Gh 26 (cagE-negative), was non-typeable by AP-PCR. Of the strains isolated in Ghana, 15 are AP-PCR type 1, 13 type 2, and 12 type 3. Of the strains isolated in Sweden, two are AP-PCR type 1, two are type 2 and nine type 3. Moreover, A. actinomycetemcomitans strains HK908 (AP-PCR type 2), originally referred to as non-serotypeable (Haubek et al., 2007), but later confirmed as serotype b (Johansson et al., 2017), and HK912 (AP-PCR type 1; serotype b) (Haubek et al., 2007) were used as references. All strains were cultured on blood agar plates (5% defibrinated horse blood, 5 mg hemin/L, 10 mg vitamin K/L, Columbia agar base) and incubated in air supplemented with 5% CO2, at 37°C.
Multilocus Sequence Typing (MLST) and Evolutionary Analyses
The gene fragments used for MLST in this study were originally selected from the A. actinomycetemcomitans serotype b, JP2 genotype strain, named HK1651 (Haubek et al., 2006), which were utilized for the analysis of JP2 genotype strains (Haubek et al., 2007). In the present study, fragments of genes encoding four housekeeping enzymes were selected for MLST: recA (GenBank accession EF142768) encoding the RecA protein, adk (EF142164) encoding adenylate kinase, frdB (EF142336) encoding fumarate reductase, and atpG (EF142218) encoding the gamma subunit of ATP synthase F1. In addition, two fragments of the hemoglobin-binding protein pseudogene hbpA (hbpA-1, and hbpA-2; EF142408 and EF142489), and one fragment of the transferring-binding protein pseudogene, tbpA (EF142817). Templates for PCR were obtained by boiling a loopful of A. actinomycetemcomitans colonies in water. PCR amplification and DNA sequencing were performed as previously described (Haubek et al., 2007). The MLST sequence types (ST) of the 56 non-JP2 genotype strains were defined based on single nucleotide polymorphisms (SNPs) within the seven DNA fragments. A cluster analysis of the 56 A. actinomycetemcomitans strains was performed based on the concatenated sequences of the seven DNA fragments in the order adk, atpG, frdB, recA, hbpA-1, hbpA-2, and tbpA. Evolutionary analyses were conducted by using the Minimum Evolution algorithm in MEGA, version 7.21 (Tamura et al., 2013). A total of 3176 nucleotide positions were used in the final dataset.
Ethical Considerations
Ethical clearance for this work was obtained from the Noguchi Memorial Institute for Medical Research, University of Ghana (IRB 000 1276), and from the local ethical committee of Umeå University, Sweden (Dnr 2010-188-31M).
Results
Identification of Polymorphic Sites and SNPs
The MLST analysis of a total of 56 non-JP2 genotype strains of A. actinomycetemcomitans serotype b, including the reference strains HK908 and HK912, was based on four house-keeping genes and three additional gene fragments, as described in Materials and Methods. When using the JP2 genotype strain, HK1651 as a reference for the comparisons, 14 polymorphic sites could be identified (Table 1). However, four polymorphic sites, located in the atpG (at nucleotide [nt] position 231), hbpA-1 (at nt 44), and in the hbpA-2 (at nt 6 and nt 214) gene fragments, respectively, were unique for HK1651, and there was no variation among the 56 non-JP2 genotype strains at these sites. In these strains, on the other hand, six polymorphic sites were identified in one strain each: adk (at nt 107), atpG (at nt 459 and 479), frdB (at nt 143), recA (at nt 111), and hbpA-2 (at nt 123). In all of the 56 non-JP2 genotype strains, a total of 290 SNPs were detected at the ten polymorphic sites, representing ≈0.1% of the total number of nucleotides of the sequenced gene fragments.
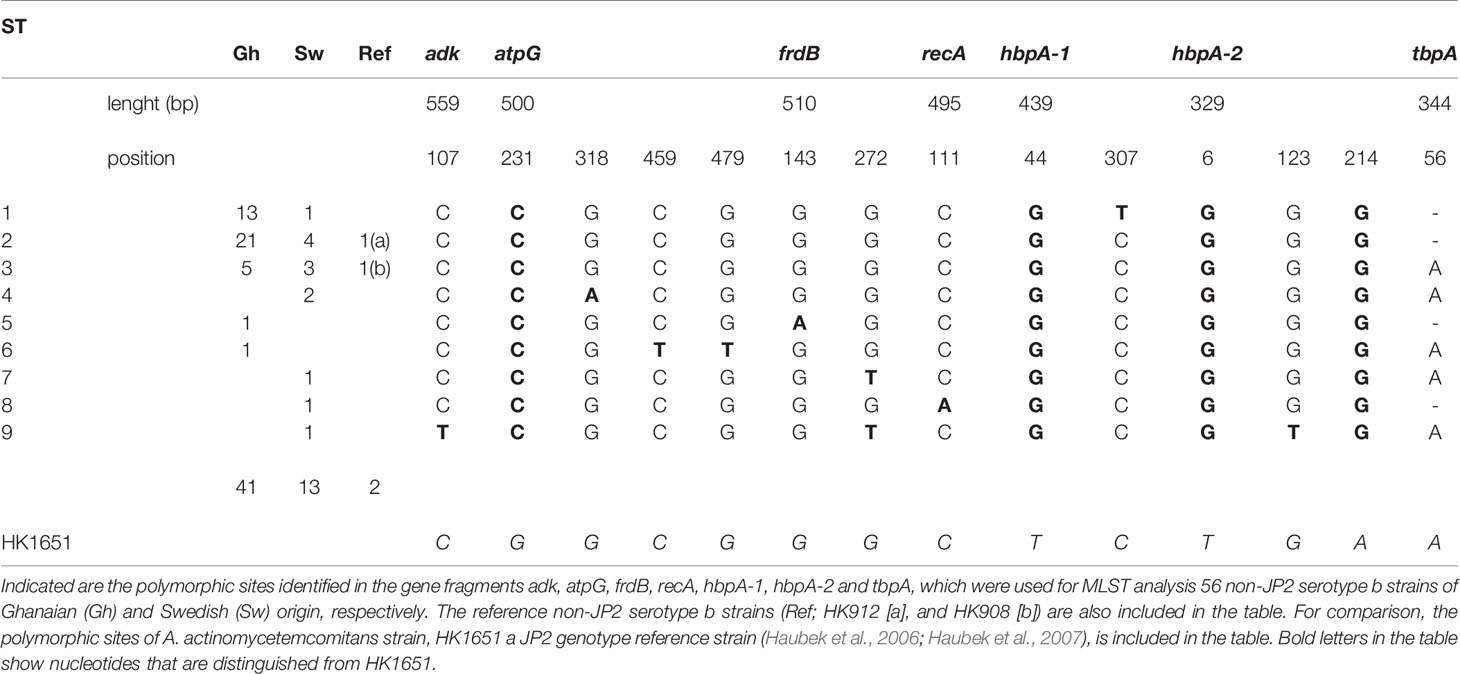
Table 1 The polymorphic sites of the nine sequence types (ST) identified by MLST analysis of 56 non-JP2 genotype, serotype b A. actinomycetemcomitans strains.
Identification of Sequence Types
Based on the MLST scheme used, nine sequence types (ST) could be identified among the 56 non-JP2 genotype strains of A. actinomycetemcomitans serotype b, and which in this work were denoted ST 1 to ST 9 (Table 2). The two non-JP2 genotype serotype b reference strains, HK908 and HK912, were confirmed to belong to ST 3 and ST 2, respectively. The overall most common sequence types among the 56 strains were ST 1 (25%), and ST 2 (46.3%). The number of SNPs among the different ST varied between four and seven (Table 1).
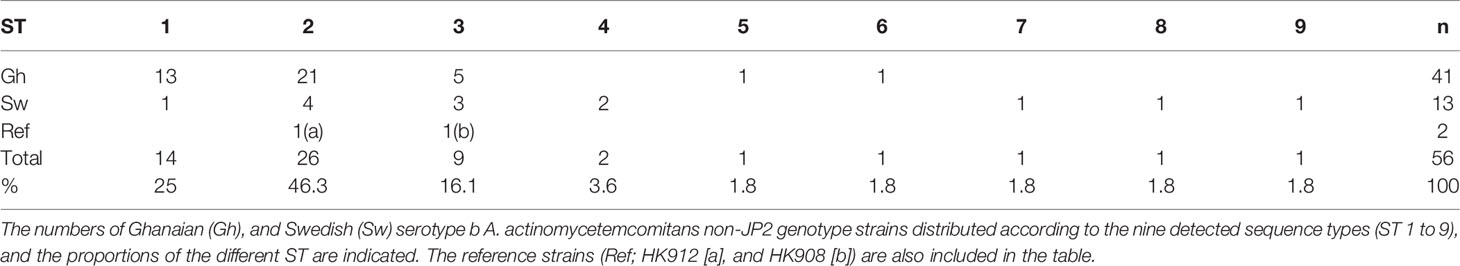
Table 2 Sequence types (ST) of the 56 non-JP2 serotype b A. actinomycetemcomitans strains.
Distribution of Sequence Types Among the Strains Isolated in Ghana and Sweden and Cluster Analysis
As indicated in Table 2, a large portion of the non-JP2 genotype strains of A. actinomycetemcomitans of serotype b isolated in Ghana and Sweden, respectively, were distributed in ST 1 to 3, which were the only sequence types identified in both countries. This supports that geographic dissemination of the non-JP2 genotype of A. actinomycetemcomitans serotype b may exist. In both collections of non-JP2 genotype serotype b strains, ST2 was the most common, representing 51.2% of the Ghanaian and 30.8% of the Swedish strains, respectively. The second most frequent sequence type, ST 1, was more common among the strains of Ghanaian (n=13) relative to those of Swedish origin (n=1). Moreover, whereas ST 5 and 6 were only identified among the Ghanaian strains, ST 4, 7, 8, and 9 were unique for the strains isolated in Sweden (Table 2). Two major clusters could be identified based on the presence and absence of a deletion of an “A” at nt position 56 in tbpA, respectively (Figure 1). The cluster lacking this deletion, referred to as cluster A, contained 14 strains, which were distributed in ST 3, 4, 6, 7, and 9. Cluster B contained 42 strains with the deletion in tbpA, i.e., these strains exhibited a deleted “A” at nt position 56 in tbpA (ST 1, 2, 5 and 8). Among the 41 Ghanaian strains, 35 (85%) were distributed in cluster B, consistent with a rather homogeneous population of non-JP2 genotype strains of serotype b. In contrast, the strains of Swedish origin appeared to be more heterogeneous, with seven (54%) belonging to cluster A, and the remaining 6 (46%) to cluster B. This difference may reflect that there is a diversity in the genetic composition between the strains of Ghanaian and Swedish origin. However, as shown in the phylogenetic tree (Figure 2), cluster analysis, using the seven concatenated gene sequences revealed a close genetic relationship between essentially all 56 strains regardless of country of origin. We observed though that all 14 ST 1 strains in cluster B were represented in a separate monophyletic branch, and that strain Sw 10 (ST 9) appeared more distantly related.
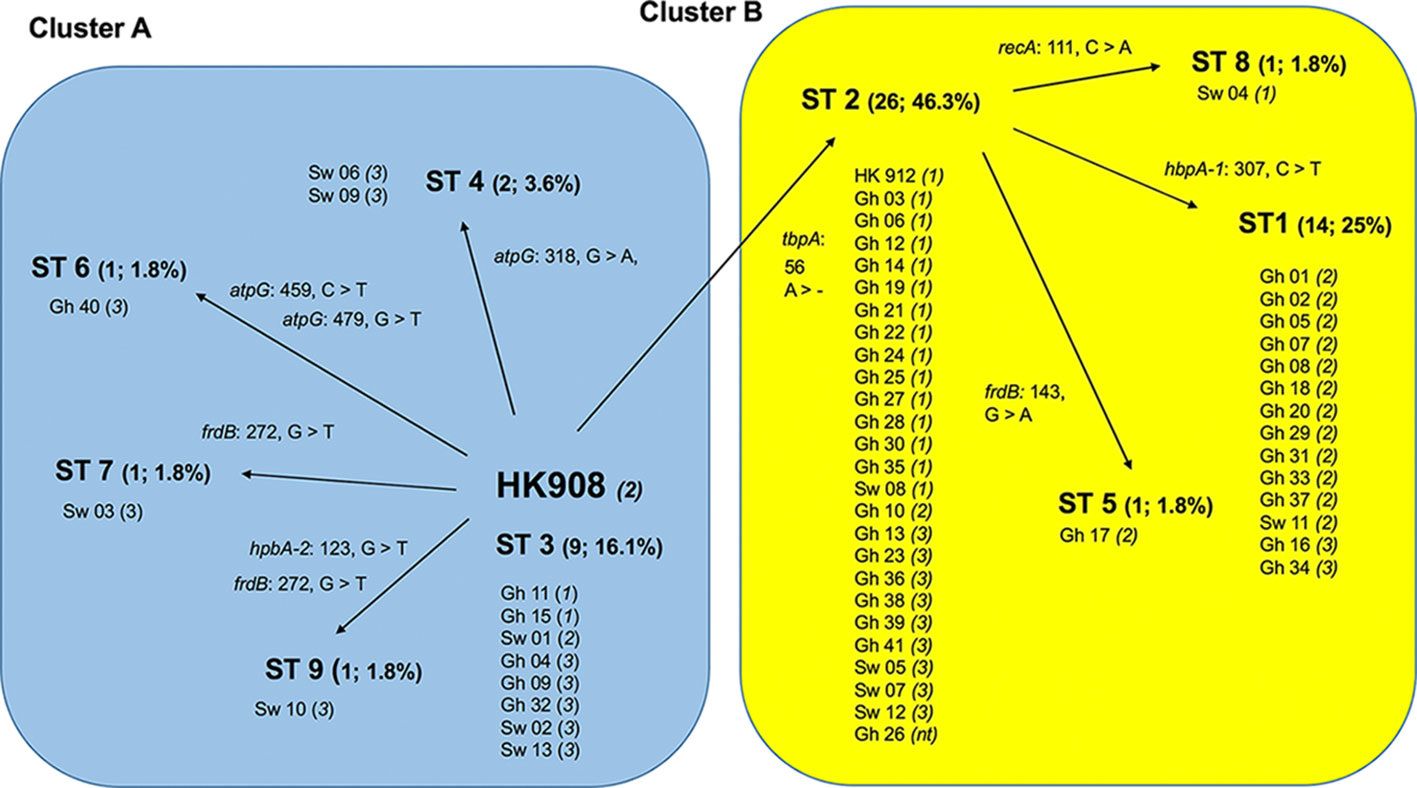
Figure 1 Schematic illustration of the separation of 56 non-JP2 genotype strains of A. actinomycetemcomitans, serotype b strains into two clusters by MLST. Cluster A (blue background), and B (yellow), respectively, were separated based on the nine identified sequence types (ST 1-9). The numbers and proportions of each ST out of the 56 strains are indicated. All ST distributed into Cluster B carry the deletion of an “A” at nucleotide position 56 in tbpA (tbpA 56 >-). The AP-PCR-type (1-3, or non-typeable [nt]) is indicated within parentheses for each strain. Gh, Ghanaian strains; Sw, strains of Swedish origin. The reference strains HK908 and HK912 are included in the diagram.
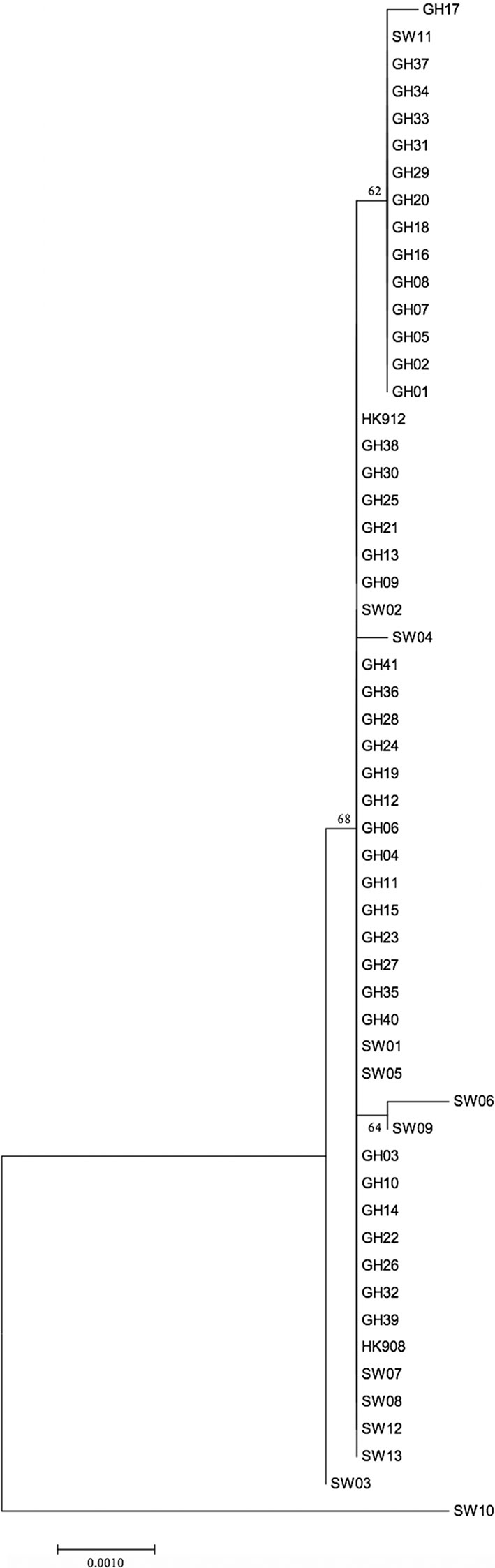
Figure 2 Phylogenetic relationship of 56 non-JP2 genotype A. actinomycetemcomitans strains of serotype b, based on MLST. The phylogenetic tree with the highest log likelihood is shown. The percentages of trees, in which the associated strains clustered together, is shown next to the branches. The tree is drawn to scale, with branch lengths measured in the number of substitutions per site. The bar indicates genetic distance. GH, Ghanaian strains; SW, Swedish strains.
Correlation Between MLST-Based Sequence Types and AP-PCR Genotype
The 56 non-JP2 genotype strains of A. actinomycetemcomitans, serotype b strains were earlier assessed by AP-PCR, and were distributed into AP-PCR types 1, 2 and 3, as described in Materials and Methods. The AP-PCR type of each of the nine MLST-based ST found among the Ghanaian and Swedish strains together, including the reference strains HK908 and HK912, is indicated in Table 3. Among the 18 AP-PCR type 1 (and cagE+) strains, a large majority, i.e., 14 (77.8%) were found to belong to ST 2. However, ST 2 did not exhibit any unique SNP at the ten polymorphic sites (Table 1). Of the 16 AP-PCR type 2 strains, the majority (12; 75%) belonged to ST 1. Thus, the polymorphic site at nt 307 in hbpA-1 (Table 1), unique for ST 1 strains, could be considered rather strongly associated with AP-PCR type 2. The 21 AP-PCR type 3 strains, were as expected a less homogeneous group, and were distributed into ST 1, 2, 3, 4, 6, 7, and 9. Based on these observations together, we could conclude that the AP-PCR genotypes to some extent do cluster into specific MLST-defined ST, but that cagE-genotyping presently appears to be a stronger, and more easily implemented tool to diagnose carriage of virulent non-JP2 genotype strains of A. actinomycetemcomitans serotype b.
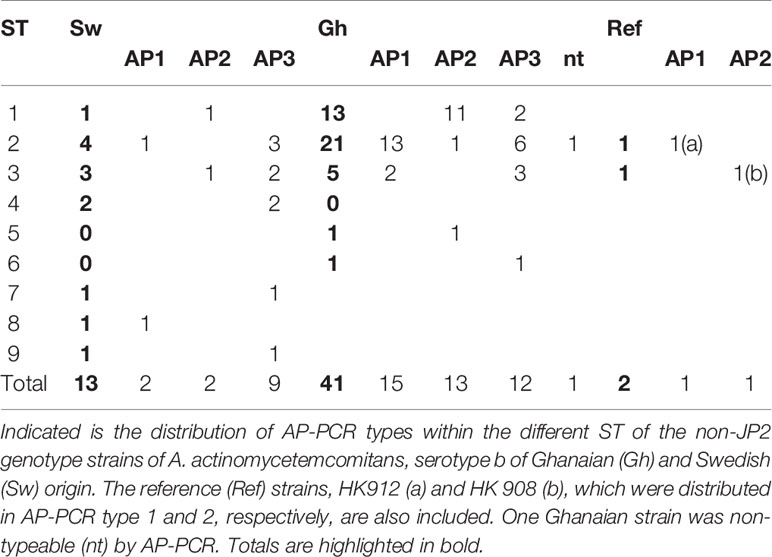
Table 3 AP-PCR types of the 56 non-JP2 serotype b A. actinomycetemcomitans strains.
Discussion
In this work, we have used MLST to investigate the genetic diversity of 56 non-JP2 genotype strains of A. actinomycetemcomitans serotype b, with 41 strains originating from Ghana and 13 from Sweden.
Interestingly, serotype b includes a genetic variant that can be found both among JP2 genotype, and among some non-JP2 genotype strains. We have earlier demonstrated that these strains belong to the same AP-PCR type (i.e., AP-PCR type 1) (Johansson et al., 2017). While JP2 genotype strains of A. actinomycetemcomitans previously have been characterized by MLST, corresponding studies of non-JP2 strains of serotype b have been scarcely reported on (Haubek et al., 2007). Based on the MLST analysis of the 56 non-JP2 genotype serotype b strains, we suggest that the overall mutation rate of this genotype is rather low. Comparing the nine identified MLST sequence types with their corresponding AP-PCR genotype, we found both similarities and discrepancies, as the strains were distributed in different clusters based on the two genotyping methods.
In this study, although the mutation rate within the non-JP2 genotype strains appeared to be low as evidenced by MLST, discrepancies between the strains of Ghanaian and Swedish origin were detected. This was emphasized by the five unique SNPs among the Ghanaian strains, and seven among the strains isolated from individuals living in Sweden, despite that the number of assessed Swedish strains was more than three times lower. This indicates that the Swedish strains may represent a more diverse group. The reason for this could probably be that the Ghanaian collection of strains was sampled from a more homogeneous group of individuals with regards to age and geographic localization (Höglund Åberg et al., 2012). The notion that A. actinomycetemcomitans serotype b that colonizes the Swedish population, are more prone to mutations is another possible explanation.
A. actinomycetemcomitans strain HK908 is one of few non-JP2 genotype serotype b strains that has been analyzed previously by MLST (Haubek et al., 2007). This strain was in that study described as non-serotypeable. As HK908 in the study by Haubek and coworkers (Haubek et al., 2007) was used to illustrate the evolutionary stage, where non-JP2 genotype A. actinomycetemcomitans serotype b evolved into the JP2 genotype, we chose to include this strain also in the present study, after confirmation that it did in fact belong to serotype b (Johansson et al., 2017). For further comparisons, also the non-JP2 genotype serotype b strain, HK 912 was included as a reference. In the study by Haubek and coworkers (Haubek et al., 2007), HK908 was demonstrated to represent a breaking point, dividing JP2 and non-JP2 genotype strains of A. actinomycetemcomitans of serotype b into two different clusters. In this work, HK908 was instead used to illustrate clustering among non-JP2 genotype strains. As the deletion of an “A” at nt position 56 in tbpA was observed only in non-JP2 genotype serotype b strains in the previous study (Haubek et al., 2007), we here used the presence and the absence of this deletion as a means to divide the 56 non-JP2 genotype serotype b strains into two major clusters, A and B, respectively. This led to the observation that strains of Ghanaian and Swedish origin were differentially distributed in these clusters, again indicating that there is a genetic diversity between the two collections of non-JP2 genotype strains of A. actinomycetemcomitans serotype b from Ghana and Sweden, respectively.
Different types of PCR-based methods for the characterization of bacterial genomes have been developed during the last decades (van Belkum, 1994; Jolley and Maiden, 2014; Bonk et al., 2018; Wade and Prosdocimi, 2020). However, similar results will not necessarily be obtained, when bacterial collections are characterized with other PCR-based methods (Tsang et al., 2017). When we earlier characterized non-JP2 genotype strains of A. actinomycetemcomitans, serotype b by AP-PCR, we found more than 10 different banding patterns. The strains were grouped into AP-PCR types 1, 2 and others (referred to as AP-PCR type 3 in this work) (Claesson et al., 2017; Johansson et al., 2019). Among serotype b, strains of AP-PCR type 1 are in general more leukotoxic than the other AP-PCR types, and they are more frequently associated with periodontitis affecting young individuals (Dogan et al., 1999a; Höglund Åberg et al., 2014). Thus, albeit AP-PCR–based analyses of A. actinomycetemcomitans mainly reflects the general genetic diversity within this bacterial species, this methodology can be valuable for diagnostic purposes, to identify highly leukotoxic strains of serotype b. However, evidently AP-PCR is also a time-consuming method, which is associated with difficulties in standardization (Meunier and Grimont, 1993; Power, 1996; Tyler et al., 1997; Suzuki et al., 2003).
As evidenced from our present study, the MLST scheme implemented appeared not to be a highly discriminatory method to identify AP-PCR type 1, as strains of this AP-PCR type could be found within three different ST-types (ST 2, ST 3 and ST 88). Interestingly, AP-PCR type 1 A. actinomycetemcomitans strains of serotype b, isolated from both Ghana and Sweden, regardless being of JP2 or non-JP2 genotype, carry a gene that with very few known exceptions is present only in this AP-PCR type, i.e., cagE (Johansson et al., 2017; Johansson et al., 2019). As discussed earlier (Johansson et al., 2017; Johansson et al., 2019), the cagE gene product per se is most likely not responsible for the increased leukotoxicity of AP-PCR type 1 strains. However, to identify whether carriers are infected with a more virulent A. actinomycetemcomitans serotype b, i.e., AP-PCR type 1, which includes both JP2 and non-JP2 genotypes, detection of the cagE gene by PCR-based methodology appears to represent a more efficient tool than MLST.
Conclusions
Although based on a limited number of assessed A. actinomycetemcomitans strains in this work, especially those collected from carriers in Sweden, there is a clear indication of a geographic dissemination across continents of non-JP2 genotype A. actinomycetemcomitans serotype b. Detection of the cagE gene is probably the currently known, most straightforward diagnostic procedure for detection of patients carrying the virulent AP-PCR type 1 of A. actinomycetemcomitans, serotype b. The cagE gene as a marker can be used to identify both JP2 and non-JP2 genotype strains of A. actinomycetemcomitans serotype b of this AP-PCR type, with PCR or qPCR, and cagE-specific oligonucleotide primers.
Data Availability Statement
The original contributions presented in the study are included in the article/Supplementary Material. Further inquiries can be directed to the corresponding author.
Author Contributions
Conceptualization, RC, AJ, and JO. Methodology, RC, AJ, AE, and JO. Validation, RC, AJ, CH, AE, and JO. Formal analysis, RC, AJ, and JO. Investigation, RC, AJ, and JO. Resources RC, AJ, CH, DH, and JO. Data curation, RC, AJ, and JO. Writing—original draft preparation, RC, AJ, and JO. Writing—review and editing, RC, AJ, CH, AE, DH, and JO. Visualization, RC, AJ, DH, and JO. Supervision, AJ, RC, and JO. Project administration, RC, AJ, and JO. funding acquisition, AJ and JO. All authors contributed to the article and approved the submitted version.
Funding
This work was supported by TUA grants from the Region of Västerbotten, Sweden (AJ; 7003193 and JO; 7002667), and from Insamlingsstiftelsen, Medical Faculty, Umeå University, Sweden.
Conflict of Interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Publisher’s Note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Acknowledgments
We are grateful to Chrissie Roth and Ewa Strömkvist-Engbo for valuable technical assistance.
Supplementary Material
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fcimb.2021.769671/full#supplementary-material
References
Asikainen, S., Chen, C., Slots, J. (1995). Actinobacillus actinomycetemcomitans Genotypes in Relation to Serotypes and Periodontal Status. Oral. Microbiol. Immunol. 10 (2), 65–68.
Asikainen, S., Karched, M. (2008). “Molecular Techniques in Oral Microbial Taxonomy, Identification and Typing,” in Molecular Oral Microbiology. Ed. Rogers, A. H. (Adelaide: Caister Academic Press).
Bonk, F., Popp, D., Harms, H., Centler, F. (2018). PCR-Based Quantification of Taxa-Specific Abundances in Microbial Communities: Quantifying and Avoiding Common Pitfalls. J. Microbiol. Methods 153, 139–147. doi: 10.1016/j.mimet.2018.09.015
Brogan, J. M., Lally, E. T., Poulsen, K., Kilian, M., Demuth, D. R. (1994). Regulation of Actinobacillus actinomycetemcomitans Leukotoxin Expression: Analysis of the Promoter Regions of Leukotoxic and Minimally Leukotoxic Strains. Infect. Immun. 62 (2), 501–508.
Bueno, L. C., Mayer, M. P., DiRienzo, J. M. (1998). Relationship Between Conversion of Localized Juvenile Periodontitis-Susceptible Children From Health to Disease and Actinobacillus actinomycetemcomitans Leukotoxin Promoter Structure. J. Periodontol. 69 (9), 998–1007. doi: 10.1902/jop.1998.69.9.998
Burgess, D., Huang, H., Harrison, P., Aukhil, I., Shaddox, L. (2017). Aggregatibacter actinomycetemcomitans in African Americans With Localized Aggressive Periodontitis. JDR Clin. Trans. Res. 2 (3), 249–257. doi: 10.1177/2380084417695543
Claesson, R., Chiang, H. M., Lindholm, M., Höglund Åberg, C., Haubek, D., Johansson, A., et al. (2020). Characterization of Aggregatibacter actinomycetemcomitans Serotype B Strains With Five Different, Including Two Novel, Leukotoxin Promoter Structures. Vaccines (Basel) 8 (3), 398. doi: 10.3390/vaccines8030398
Claesson, R., Gudmundson, J., Höglund Åberg, C., Haubek, D., Johansson, A. (2015). Detection of a 640-Bp Deletion in the Aggregatibacter actinomycetemcomitans Leukotoxin Promoter Region in Isolates From an Adolescent of Ethiopian Origin. J. Oral. Microbiol. 7:26974. doi: 10.3402/jom.v7.26974
Claesson, R., Höglund-Åberg, C., Haubek, D., Johansson, A. (2017). Age-Related Prevalence and Characteristics of Aggregatibacter actinomycetemcomitans in Periodontitis Patients Living in Sweden. J. Oral. Microbiol. 9 (1):1334504. doi: 10.1080/20002297.2017.1334504
Dogan, B., Asikainen, S., Jousimies-Somer, H. (1999a). Evaluation of Two Commercial Kits and Arbitrarily Primed PCR for Identification and Differentiation of Actinobacillus actinomycetemcomitans, Haemophilus aphrophilus, and Haemophilus paraphrophilus. J. Clin. Microbiol. 37 (3), 742–747.
Dogan, B., Saarela, M., Asikainen, S. (1999b). Genotyping of Actinobacillus actinomycetemcomitans Serotype D Isolates Based on Polymerase Chain Reaction. Oral. Microbiol. Immunol. 14 (6), 387–390.
Fine, D. H., Schreiner, H., Velusamy, S. K. (2020). Aggregatibacter, a Low Abundance Pathobiont That Influences Biogeography, Microbial Dysbiosis, and Host Defense Capabilities in Periodontitis: The History of a Bug, and Localization of Disease. Pathogens 9 (3), 179. doi: 10.3390/pathogens9030179
Haraszthy, V. I., Hariharan, G., Tinoco, E. M., Cortelli, J. R., Lally, E. T., Davis, E., et al. (2000). Evidence for the Role of Highly Leukotoxic Actinobacillus actinomycetemcomitans in the Pathogenesis of Localized Juvenile and Other Forms of Early-Onset Periodontitis. J. Periodontol. 71 (6), 912–922. doi: 10.1902/jop.2000.71.6.912
Haubek, D. (2010). The Highly Leukotoxic JP2 Clone of Aggregatibacter actinomycetemcomitans: Evolutionary Aspects, Epidemiology and Etiological Role in Aggressive Periodontitis. APMIS Suppl. (130), 1–53. doi: 10.1111/j.1600-0463.2010.02665.x
Haubek, D., Ennibi, O. K., Poulsen, K., Vaeth, M., Poulsen, S., Kilian, M. (2008). Risk of Aggressive Periodontitis in Adolescent Carriers of the JP2 Clone of Aggregatibacter (Actinobacillus) Actinomycetemcomitans in Morocco: A Prospective Longitudinal Cohort Study. Lancet 371 (9608), 237–242. doi: 10.1016/S0140-6736(08)60135-X
Haubek, D., Havemose-Poulsen, A., Westergaard, J. (2006). Aggressive Periodontitis in a 16-Year-Old Ghanaian Adolescent, the Original Source of Actinobacillus actinomycetemcomitans Strain HK1651 - a 10-Year Follow Up. Int. J. Paediatr. Dent. 16 (5), 370–375. doi: 10.1111/j.1365-263X.2006.00735.x
Haubek, D., Poulsen, K., Kilian, M. (2007). Microevolution and Patterns of Dissemination of the JP2 Clone of Aggregatibacter (Actinobacillus) actinomycetemcomitans. Infect. Immun. 75 (6), 3080–3088. doi: 10.1128/IAI.01734-06
Henderson, B., Ward, J. M., Ready, D. (2010). Aggregatibacter (Actinobacillus) actinomycetemcomitans: A Triple A* Periodontopathogen? Periodontol. 2000 54 (1), 78–105.
He, T., Nishihara, T., Demuth, D. R., Ishikawa, I. (1999). A Novel Insertion Sequence Increases the Expression of Leukotoxicity in Actinobacillus actinomycetemcomitans Clinical Isolates. J. Periodontol. 70 (11), 1261–1268. doi: 10.1902/jop.1999.70.11.1261
Höglund Åberg, C., Haubek, D., Kwamin, F., Johansson, A., Claesson, R. (2014). Leukotoxic Activity of Aggregatibacter actinomycetemcomitans and Periodontal Attachment Loss. PloS One 9 (8), e104095. doi: 10.1371/journal.pone.0104095
Höglund Åberg, C., Kwamin, F., Claesson, R., Johansson, A., Haubek, D. (2012). Presence of JP2 and non-JP2 Genotypes of Aggregatibacter actinomycetemcomitans and Attachment Loss in Adolescents in Ghana. J. Periodontol. 83 (12), 1520–1528. doi: 10.1902/jop.2012.110699
Johansson, A., Claesson, R., Höglund Åberg, C., Haubek, D., Lindholm, M., Jasim, S., et al. (2019). Genetic Profiling of Aggregatibacter actinomycetemcomitans Serotype B Isolated From Periodontitis Patients Living in Sweden. Pathogens 8 (3), 153. doi: 10.3390/pathogens8030153
Johansson, A., Claesson, R., Höglund Åberg, C., Haubek, D., Oscarsson, J. (2017). The cagE Gene Sequence as a Diagnostic Marker to Identify JP2 and non-JP2 Highly Leukotoxic Aggregatibacter actinomycetemcomitans Serotype B Strains. J. Periodontal Res. 52 (5), 903–912. doi: 10.1111/jre.12462
Jolley, K. A., Maiden, M. C. (2014). Using Multilocus Sequence Typing to Study Bacterial Variation: Prospects in the Genomic Era. Future Microbiol. 9 (5), 623–630. doi: 10.2217/fmb.14.24
Kittichotirat, W., Bumgarner, R. E., Chen, C. (2016). Evolutionary Divergence ofAggregatibacter actinomycetemcomitans. J. Dent. Res. 95 (1), 94–101. doi: 10.1177/0022034515608163
Kolodrubetz, D., Dailey, T., Ebersole, J., Kraig, E. (1989). Cloning and Expression of the Leukotoxin Gene From Actinobacillus actinomycetemcomitans. Infect. Immun. 57 (5), 1465–1469. doi: 10.1128/IAI.57.5.1465-1469.1989
Könönen, E., Muller, H. P. (2014). Microbiology of Aggressive Periodontitis. Periodontol. 2000 65 (1), 46–78. doi: 10.1111/prd.12016
Meunier, J. R., Grimont, P. A. (1993). Factors Affecting Reproducibility of Random Amplified Polymorphic DNA Fingerprinting. Res. Microbiol. 144 (5), 373–379. doi: 10.1016/0923-2508(93)90194-7
Power, E. G. (1996). RAPD Typing in Microbiology–a Technical Review. J. Hosp. Infect. 34 (4), 247–265. doi: 10.1016/s0195-6701(96)90106-1
Rylev, M., Kilian, M. (2008). Prevalence and Distribution of Principal Periodontal Pathogens Worldwide. J. Clin. Periodontol. 35 (8 Suppl), 346–361. doi: 10.1111/j.1600-051X.2008.01280.x
Spitznagel, J., Jr., Kraig, E., Kolodrubetz, D. (1991). Regulation of Leukotoxin in Leukotoxic and Nonleukotoxic Strains of Actinobacillus actinomycetemcomitans. Infect. Immun. 59 (4), 1394–1401. doi: 10.1128/IAI.59.4.1394-1401.1991
Suzuki, K., Ohnami, S., Tanabe, C., Sasaki, H., Yasuda, J., Katai, H., et al. (2003). The Genomic Damage Estimated by Arbitrarily Primed PCR DNA Fingerprinting Is Useful for the Prognosis of Gastric Cancer. Gastroenterology 125 (5), 1330–1340. doi: 10.1016/j.gastro.2003.07.006
Tamura, K., Stecher, G., Peterson, D., Filipski, A., Kumar, S. (2013). MEGA6: Molecular Evolutionary Genetics Analysis Version 6.0. Mol. Biol. Evol. 30 (12), 2725–2729. doi: 10.1093/molbev/mst197
Tsai, C. C., Ho, Y. P., Chou, Y. S., Ho, K. Y., Wu, Y. M., Lin, Y. C. (2018). Aggregatibacter (Actinobacillus) actimycetemcomitans Leukotoxin and Human Periodontitis - A Historic Review With Emphasis on JP2. Kaohsiung J. Med. Sci. 34 (4), 186–193. doi: 10.1016/j.kjms.2018.01.014
Tsang, A. K. L., Lee, H. H., Yiu, S. M., Lau, S. K. P., Woo, P. C. Y. (2017). Failure of Phylogeny Inferred From Multilocus Sequence Typing to Represent Bacterial Phylogeny. Sci. Rep. 7 (1), 4536. doi: 10.1038/s41598-017-04707-4
Tyler, K. D., Wang, G., Tyler, S. D., Johnson, W. M. (1997). Factors Affecting Reliability and Reproducibility of Amplification-Based DNA Fingerprinting of Representative Bacterial Pathogens. J. Clin. Microbiol. 35 (2), 339–346. doi: 10.1128/jcm.35.2.339-346.1997
van Belkum, A. (1994). DNA Fingerprinting of Medically Important Microorganisms by Use of PCR. Clin. Microbiol. Rev. 7 (2), 174–184. doi: 10.1128/CMR.7.2.174
Keywords: Aggregatibacter actinomycetemcomitans, multilocus sequence typing (MLST), non-JP2, serotype b, cagE, evolutionary analysis, genomic analysis
Citation: Claesson R, Johansson A, Höglund Åberg C, Esberg A, Haubek D and Oscarsson J (2021) Multilocus Sequence Typing of Non-JP2 Serotype b Aggregatibacter actinomycetemcomitans Strains of Ghanaian and Swedish Origin. Front. Cell. Infect. Microbiol. 11:769671. doi: 10.3389/fcimb.2021.769671
Received: 02 September 2021; Accepted: 26 November 2021;
Published: 14 December 2021.
Edited by:
Gunnar Dahlen, University of Gothenburg, SwedenReviewed by:
Senthil Kumar Velusamy, Rutgers Biomedical and Health Sciences, United StatesJaroslav Hrabak, Charles University, Czechia
Copyright © 2021 Claesson, Johansson, Höglund Åberg, Esberg, Haubek and Oscarsson. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Jan Oscarsson, jan.oscarsson@umu.se