

Abstract
Grassland soils represent a significant potential pool for the sequestering of atmospheric CO2; however, the magnitude of this pool depends to a certain extent on the grazing management of the grassland. Whether different herbivore species and combinations (herbivore assemblage) influence the soil organic carbon (SOC) in various grasslands remains unclear. Here we have investigated the impacts of herbivore assemblages (NG, no grazing; SG, sheep grazing; CG, cattle grazing; and MG, mixed grazing of sheep and cattle) under moderate grazing intensity on a shallow layer of SOC storage across the three different grassland types (meadow steppe, typical steppe, and desert steppe) in the temperate steppes of Eastern Eurasia. We found that herbivore-species-specific effects on SOC storage depended on the grassland type. In mesic grasslands (i.e. meadow steppe and typical steppe), CG enhanced SOC storage, but there were no effects from SG. In contrast, SG in the xeric grassland (i.e. desert steppe) dramatically increased SOC storage while no effects were observed for CG. Importantly, MG of sheep and cattle consistently increased SOC storage across all grassland types. We suggest that adopting herbivore-type-specific stewardship in different grasslands could assist with the enhancement of ecosystem C functionality and services; large herbivores and small herbivores are suitable for grazing in mesic grasslands and xeric grasslands, respectively. Moreover, MG with diverse herbivores may be the optimal moderate grazing mode for soil C sequestration in most grasslands of northern China.
Export citation and abstract BibTeX RIS

Original content from this work may be used under the terms of the Creative Commons Attribution 4.0 license. Any further distribution of this work must maintain attribution to the author(s) and the title of the work, journal citation and DOI.
1. Introduction
Soil organic carbon (SOC) in grasslands represents a potential C pool, where the C pool in the shallow layer accounts for more than 60% of the total C pool in the top 1 m depth. However, it is vulnerable to loss due to intensive livestock activities (Piñeiro et al 2009, Yang et al 2010, McSherry and Ritchie 2013). Most indigenous communities have for centuries managed rangelands for livestock grazing, which may affect the ability of shallow layer grassland soils to store carbon (Vallentine 2000). A shift to strategic grazing management in grasslands can promote soil C sequestration, and potentially offset a portion of previous C emissions (Bagchi and Ritchie 2010, Wilson et al 2018). However, there are ongoing debates about how to optimize herbivore management to boost soil C sequestration (Conant et al 2001). Remarkably, this lack of consensus affects assessments and predictions of how grazing patterns will influence soil C pools and restore healthy ecosystem functioning in grasslands in the face of climate change (Eldridge et al 2016, Derner et al 2018).
Herbivores influence soil C sequestration via two separate but related processes (figure 1). First, the foraging behavior of herbivores affects plant community composition, as well as the quantity and quality of plant biomass (Wang et al 2010, Charles et al 2017). Meanwhile, moderate foraging intensity by herbivores may stimulate root growth and increase root biomass, thus affecting soil C inputs (Hamilton et al 2008, Wilson et al 2018). Second, trampling, urine and dung from grazing herbivores, is associated with changes in the soil properties (Hiltbrunner et al 2012, Liu et al 2017). In turn, changes in the soil properties also influence soil microbes and their C use efficiency, and affect soil respiration and C losses (Frey et al 2001, Li et al 2018).
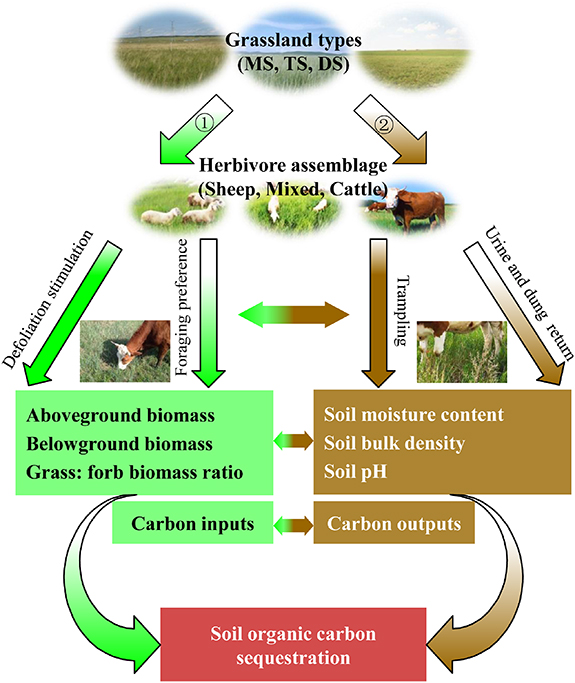
Figure 1. Schematic diagram of the effects of herbivore assemblages (sheep, cattle, mixed of sheep and cattle) on SOC (red box) when considering each of two patterns in different grassland types. Pattern 1 (green boxes and arrows, main carbon input processes) and pattern 2 (brown boxes and arrows, main carbon output processes). Curved arrows indicate the overall results of all factors. MS, meadow steppe; TS, typical steppe; and DS, desert steppe.
Download figure:
Standard image High-resolution imageDifferences in body sizes and dietary preferences among herbivore species can lead to discrepancies in diet selection, which probably alters plant community structure and composition (Bakker et al 2004, Chang et al 2018). Small herbivores tend to focus on highly nutritious forbs, which may reduce their proportion in grasslands (Edwards and Crawley 1999). In contrast, large herbivores are typically less selective and consequently may decrease the abundance of herbaceous plant species (Milchunas and Lauenroth 1993). Nevertheless, small and large herbivores within the herbivore assemblage can compete with or facilitate each other due to diet differences or overlaps (Van Der Wal et al 2000). The effects of combinations of multiple herbivores on the plant can therefore be additive or compensatory, with potentially quite idiosyncratic effects on plant community composition and diversity (Chang et al 2018). Furthermore, herbivore selective foraging accompanies interactions with trampling activity. Generally, small herbivores exhibit greater trampling frequencies and trampling steps than large herbivores. Thus, they have a stronger compaction effect on the shallow layer of soils in contrast to large herbivores (Vallentine 2000). For mixed grazing (MG) of multiple herbivores, there is cooperation and competition between browsing and trampling foraging due to the different body sizes of herbivores. Collectively, grazing regimes from different-sized herbivores might have significant impacts on plant and soil properties through foraging and trampling behaviors, which cascade on soil C inputs and outputs that subsequently influence soil C storage.
In a previous study, the effects of herbivore assemblages on soil C sequestration were examined in meadow steppe in northern China (Chang et al 2018). To the best of our knowledge, these effects have not been widely examined for different grassland types. Previous studies reported equivocal and uncertain findings of herbivore grazing effects on C sequestration under different local climates (Milchunas and Lauenroth 1993, Piñeiro et al 2010, McSherry and Ritchie 2013). A global meta-analysis indicated that precipitation could explain more variation in grazing effects on SOC for different grassland types, with the grazing effect changing from negative in mesic grasslands to positive in xeric grasslands (McSherry and Ritchie 2013). Conversely, other studies demonstrated that grazing negatively impacted SOC at lower rainfall, which was primarily due to reduced plant cover and increased bare ground (Li et al 2008, Shrestha and Stahl 2008). In addition, several studies reported that grazing increased SOC when the mean annual precipitation (MAP) was 600 mm or less (Derner and Schuman 2007, McSherry and Ritchie 2013). This was primarily attributed to a less labile C pool under drier conditions, which might reduce C turnover and increase C stock under different grazing conditions (Derner and Schuman 2007). Therefore, it is important to discover grazing-SOC patterns for different grasslands (McSherry and Ritchie 2013, Wilson et al 2018), in particular, how grazing assemblages by different herbivore types affect soil C sequestration in contrasting grasslands.
Herein, our goal was to examine the effects of herbivore assemblages on shallow layer soil C sequestration, and the applicability of herbivore types to soil C sequestration in the three grassland types under variable precipitation. Specifically, we addressed the following questions: (a) how do herbivore assemblages (SG, sheep grazing; CG, cattle grazing; and MG, mixed grazing of sheep and cattle) influence shallow layer soil C sequestration? (b) Do the effects of herbivore species on soil C sequestration depend on the grassland type? (c) Which is more important to a grazing assemblage's influence on soil C: plant functional types or soil properties?
2. Materials and methods
2.1. Study site
Our study was conducted at the main part of the Eurasian steppe, China. The entire transect spans about 1100 km, with latitudes ranging from 41° 47' to 48° 43' N, longitudes ranging from 111° 53' to 119° 55' E, and elevations spanning from 740 to 1450 m (table S1 available online at stacks.iop.org/ERL/16/114033/mmedia). The steppe range belongs to the temperate grassland and has a continental climate. Mean annual 2007–2016 temperatures along the transect range from −0.7 °C to 5.2 °C, whereas the MAP ranges from 251.1 to 354.5 mm (National Meteorological Information Center, China). Most of the precipitation is received from May to August each year. The regional transect also provides a precipitation gradient, which covers the three main community types: meadow steppe (MAP = 354.5 mm), typical steppe (MAP = 288.6 mm), and desert steppe (MAP = 251.1 mm) (table S1).
The meadow steppe located in the eastern portion of the transect is dominated by Carex pediformis C. A. Mey. and Stipa baicalensis Roshev., with the highest aboveground productivity and species richness among the three community types. The typical steppe is representative of Leymus chinensis Tzvel and S. grandis P. Smirn., with intermediate levels of productivity and the lowest species richness. The desert steppe in the western portion of the transect primarily comprises S. krylovii Roshev. and S. breviflora Griseb., with the lowest productivity and intermediate species richness. Soil types are classified as Luvisols, Phaeozems, and Arenosols for meadow steppe, typical steppe, and desert steppe, respectively (World Reference Base for Soil Resources). Detailed information on the study sites are presented in table S1.
2.2. Experimental design
Prior to the experiment, the research area with the same historical background of grazing and similar plants was enclosed to exclude livestock grazing in the meadow steppe in 2012, and in the other two grasslands in 2013 (Zhang et al 2020). The overgrazing-indicator (C4 grasses) never exceeded the critical limit of 50%. Consequently, the selected research area had not been overgrazed in recent years. Following one year of enclosure, among the three grassland types, the experiment was designed as a randomized complete block, with three blocks as replicates. Each replicate block was divided into four plots, which were assigned to four grazing treatments: NG, no grazing; SG, moderate sheep grazing; CG, moderate cattle grazing; and MG, moderate MG by sheep and cattle (figure S1). Meanwhile, each plot was enclosed with woven-wire fences (10 cm mesh size; 1.5 m high). Each plot size was calculated according to plant productivity and livestock foraging at each study site. The plot sizes of NG, SG, and CG were 0.25 ha for meadow steppe and typical steppe, and 1.32 ha for desert steppe. To ensure equal stocking rates between single grazing and MG treatments, the MG plots were twice as large as the other plots. In order to ensure that these differences in plot size (between MG and other treatments) had no direct effects on the variance of samples collected, all samples were collected at the same spatial scales and within the same total area for each of the four livestock treatments (Wang et al 2019a). Each year, grazing occurred from June to August, and grazing was maintained at a moderate grazing intensity. The numbers of sheep and cattle were standardized in accordance with their daily intakes from pre-trials (Liu et al 2017). This operation greatly minimized the differences in the stocking rate among the various treatments. For each grassland type, sheep and cattle were rotated to avoid the effects of using different animals through the nine plots (three herbivore assemblages × three replication plots) (Li et al 2019). The grazing experiment design was based on the method of rangeland carrying capacity calculation published by the agriculture industry standard NY/T 635–2015 (Ministry of Agriculture of The People's Republic of China).
2.3. Field sampling and measurements
After three to four years of moderate grazing experiments, plant and soil samples were collected in all of the experimental plots for the three grassland types in 2016. To alleviate the impacts of grassland heterogeneity on plant and soil samples, three 25 m parallel transects at 6.25 m intervals within each plot were established, and then, 50 × 50 cm quadrats along each transect at 5 m intervals were set up. For each quadrat, the abundance of each plant species was recorded, the live above-ground plant biomass was clipped at ground level, and dead parts were removed and combined with plant litter. Plant species were classified into four functional groups in terms of grasses, forbs, legumes, and sedges. The biomass of the functional groups was used to estimate the plant community composition. Since SOC is mainly concentrated in the shallow layer, and grazing-induced changes in SOC may be more likely to occur in the shallow soil layers, just the shallow samples were collected. Below-ground plant biomass and soil bulk density were evenly selected from plant quadrates to ensure the maximum coverage of the entire plot. For below-ground plant biomass, 20 cores (14 cm in diameter and 10 cm in depth) were excavated in each plot. Additionally, the soil bulk density was separated into two depths (0–5 cm, 5–10 cm), and every depth was measured with 20 cores using a soil cutting ring (volume, 50 cm3) in each plot. Further, 150 cores (2 cm in diameter and 10 cm in depth) were sampled and separated into two depths for soil property measurements in each plot. Soil samples were sieved over 2 mm mesh to separate the coarse roots and other visible debris. Samples from the same depth were mixed, packed in polyethylene bags, immediately stored in a portable refrigerator (4 °C), and transported to the laboratory for further analysis. Approximately 200 g of each soil sample was air-dried for the analysis of soil properties (e.g. SOC concentration and pH). The remaining soils were stored at 4 °C for analysis of soil moisture.
All above-ground plant biomass was oven-dried at 65 °C for 48 h to a constant weight, then weighed. Below-ground plant biomass was manually sorted from the soil, carefully washed, oven-dried at 65 °C, and weighed. SOC concentration was determined using the dichromate oxidation method (Nelson and Sommers 1982). Soil moisture was gravimetrically quantified using 10 g of fresh soil samples dried at 105 °C for 24 h to a constant weight. Soil pH was measured in a 1:2.5 (soil/water) suspension.
2.4. Data analysis
Statistical analyses were performed using R ver. 3.6.1 (R Development Core Team), significance was set at P< 0.05 level, and figures were plotted with Origin software (Version 9.0; Northampton, MA, USA). Linear mixed effect models (LMMs) were performed with grazing treatment as the fixed effect and block as the random effect to assess the impacts of grazing on SOC storage in the top two soil layers (upper layer, 0–5 cm; lower layer, 5–10 cm), plant properties (above- and below-ground plant biomass, and grass: forb biomass ratio), and soil properties (soil moisture, bulk density, and pH on upper and lower layers). The interactive effects of grazing treatment and grassland type on the above variables were also inspected. The LMMs were achieved using the function lmer in the 'lme4' package (Douglas et al 2015). Tukey tests were applied for the comparisons of grazing treatment effects, and were performed with the function glht in the 'multcomp' package (Torsten et al 2008).
The relative importance of environmental factors as explanatory variables of SOC storage was examined using LMMs with plant and edaphic factors as the fixed effects, and block as the random effect. For all models, environmental factors were added according to a fixed sequential order as plant (above- and below-ground plant biomass, and grass: forb biomass ratio), and soil (soil moisture, bulk density, pH). Before applying LMMs, we implemented a variance inflation factor test for all selected factors in order to check the multicollinearity of model terms. To reduce the effects of spurious relationships between factors, we ran the LMMs test twice. The first run was to remove the insignificant factors and select those that were significant factors for entire grasslands. Subsequently, we reran the LMMs and reported the model results of the three significant factors from the second test. The proportion of variance (R2) explained by the total fixed, random, and residual effects were estimated using the function of rsquared.lme. The total R2 of fixed effects was assigned to each factor using the pamer.fnc function from the 'LMERConvenienceFunctions' package (Tremblay et al 2013).
3. Results
According to the LMMs, herbivore assemblages and grassland types had significant effects on the shallow layer of SOC storage, plant, and soil properties (P< 0.05, table 1). The significant interactive effects of herbivore assemblages and grassland types were also detected for the majority of variables, except for soil moisture and soil bulk density (table 1).
Table 1. Statistical summary of linear mixed effect models (LMMs) evaluating the effects of herbivore assemblages, grassland types, and their interactive effects on soil organic carbon storage (SOC), above-ground plant biomass (AB), below-ground plant biomass (BB), grass: forb biomass ratio (G: F), soil moisture (SM), soil bulk density (BD), and soil pH.
| Herbivore assemblages (HA) | Grassland types (GT) | HA × GT | ||||
|---|---|---|---|---|---|---|
| Variable | F | P | F | P | F | P |
| SOC (0–5 cm) | 14.000 | <0.001 | 221.027 | <0.001 | 2.568 | 0.023 |
| SOC (5–10 cm) | 9.759 | <0.001 | 78.431 | <0.001 | 4.686 | <0.001 |
| AB | 31.061 | <0.001 | 44.461 | <0.001 | 4.640 | <0.001 |
| BB | 24.412 | <0.001 | 91.763 | <0.001 | 8.368 | <0.001 |
| G: F | 42.174 | <0.001 | 419.896 | <0.001 | 41.578 | <0.001 |
| SM (0–5 cm) | 12.132 | <0.001 | 18.280 | <0.001 | 1.346 | 0.243 |
| SM (5–10 cm) | 8.115 | <0.001 | 174.689 | <0.001 | 1.373 | 0.231 |
| BD (0–5 cm) | 15.473 | <0.001 | 101.836 | <0.001 | 0.392 | 0.883 |
| BD (5–10 cm) | 4.212 | 0.007 | 243.428 | <0.001 | 1.298 | 0.263 |
| pH (0–5 cm) | 6.971 | <0.001 | 313.019 | <0.001 | 2.976 | 0.009 |
| pH (5–10 cm) | 8.537 | <0.001 | 1167.752 | <0.001 | 5.660 | <0.001 |
3.1. SOC variability
Within the three grassland types, the effects of herbivore assemblages were diverse. In both meadow steppe and typical steppe, CG significantly increased SOC storage, and this effect was pronounced except at the lower soil layer (5–10 cm) in meadow steppe (figure 2). However, SG did not affect SOC storage in either meadow steppe or typical steppe. In contrast, in desert steppe, SG significantly increased SOC storage, while CG showed no significant effect on SOC storage (figure 2). MG significantly increased SOC storage in the three grassland types, except at the lower soil layer in meadow steppe (figure 2). The change trends of SOC concentration were similar to SOC storage in all herbivore assemblages (figure 2).
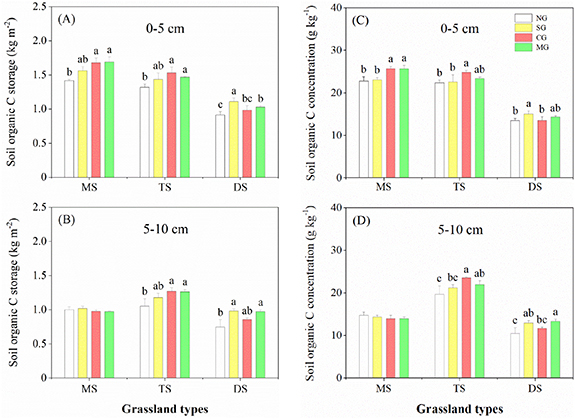
Figure 2. Effects of herbivore assemblages on upper layer SOC storage and concentration (0–5 cm) (A), (C) and lower layer SOC storage and concentration (5–10 cm) (B), (D) in various grassland types. Different letters indicate significant differences among the levels of herbivore assemblages (LMMs, P < 0.05). Error bars represent SE.
Download figure:
Standard image High-resolution image3.2. Plant communities and soil properties
For all grassland types, the plant functional type exhibited significant responses to herbivore assemblages (table 1 and figure 3). Compared to NG, above-ground plant biomass was reduced in various degrees by different herbivore assemblages among all grasslands. In contrast, below-ground plant biomass was greater under grazing treatments than NG (figure 3). In meadow steppe and typical steppe, grazing significantly increased below-ground plant biomass, except for SG in typical steppe. In desert steppe, SG and MG significantly increased below-ground plant biomass (figure 3). Moreover, SG increased the grass: forb biomass ratio 0.5–4 times in all grasslands (figure 3).
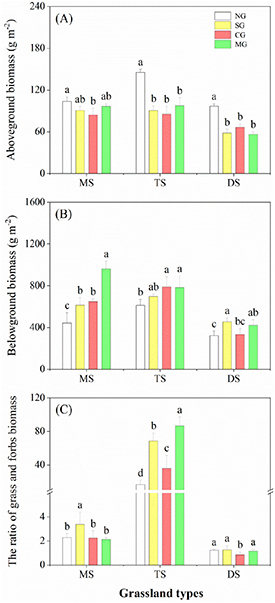
Figure 3. Above-ground plant biomass (A), below-ground plant biomass (B), and the ratio of grass and forbs biomass (C) in various grassland types for different herbivore assemblages. Different letters indicate significant differences among the levels of herbivore assemblages (LMMs, P < 0.05). Error bars represent SE.
Download figure:
Standard image High-resolution imageOverall, soil properties showed significant responses to herbivore assemblages (table 1, figures 4 and 5). Compared with NG, herbivore assemblages significantly increased soil bulk density by 4%–8%. Nonetheless, the differences among herbivore assemblages were not significant. Moreover, herbivore assemblages significantly reduced soil moisture by 2%–25% and increased soil pH by 1%–8% in different grassland types. Particularly, MG had the least impact on soil moisture compared with SG and CG (figures 4 and 5).
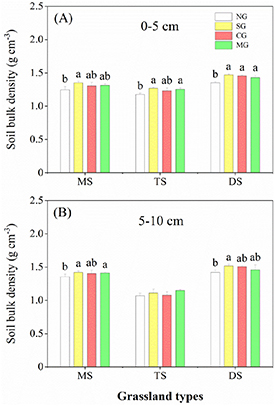
Figure 4. Effects of herbivore assemblages on upper layer soil bulk density (0–5 cm) (A) and lower layer soil bulk density (5–10 cm) (B) in various grassland types across different precipitation gradients of grassland types. Different letters indicate significant differences among the levels of herbivore assemblages (LMMs, P < 0.05). Error bars represent SE.
Download figure:
Standard image High-resolution image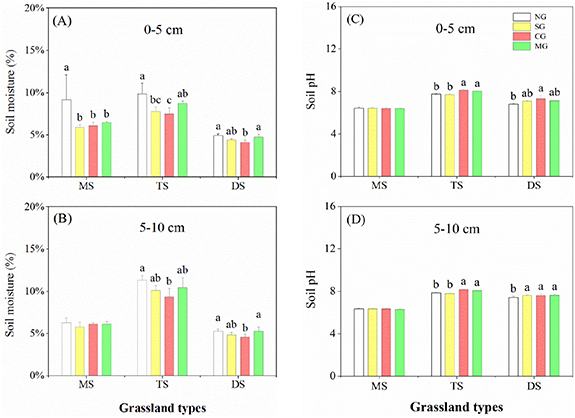
Figure 5. Effects of herbivore assemblages on upper layer soil moisture and pH (0–5 cm) (A), (C) and lower layer soil moisture and pH (5–10 cm) (B), (D) in various grassland types across different precipitation gradients of grassland types. Different letters indicate significant differences among the levels of herbivore assemblages (LMMs, P < 0.05). Error bars represent SE.
Download figure:
Standard image High-resolution image3.3. SOC storage and controlling variables
According to the results of the LMMs, the shallow layer of SOC storage was explained primarily by below-ground plant biomass, the grass: forb biomass ratio, and soil bulk density, other factors were not significant explanatory variables (table 2). In addition, below-ground plant biomass explained most of the SOC storage variability, and this effect was more evident in xeric grassland (meadow steppe: 50.9%; typical steppe: 54.6%; desert steppe: 61.3%; figure 6). Moreover, linear regression analysis also showed that SOC was positively correlated to below-ground plant biomass in all grasslands (MS: R2 = 0.51, P < 0.01; TS: R2 = 0.55, P < 0.01; DS: R2 = 0.62, P < 0.01) (figure 7).
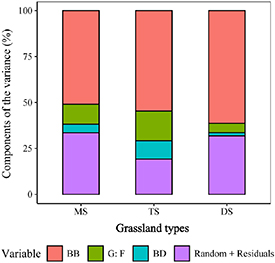
Figure 6. The percentage of variation in SOC explained by each variable in the three grassland types. BB, below-ground plant biomass; G: F, grass: forb biomass ratio; BD, soil bulk density.
Download figure:
Standard image High-resolution image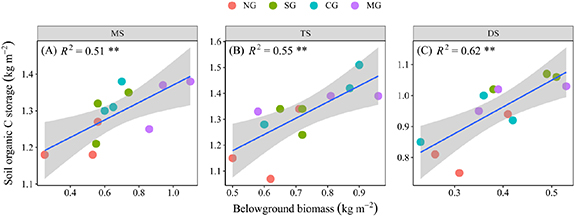
Figure 7. Relationships between SOC storage and below-ground plant biomass in various grassland types of MS (A), TS (B), and DS (C). The blue fitted lines are from ordinary least squares regression. *, ** and *** indicate significant correlation at P < 0.05, P < 0.01, P < 0.001, respectively.
Download figure:
Standard image High-resolution imageTable 2. Statistical summary of linear mixed effect models (LMMs) evaluating the effects of plant and edaphic factors on SOC in all grasslands, using all explanatory variables as fixed effects, and block as a random factor.
| Explanatory variable | df | Sum Sq | F | P |
|---|---|---|---|---|
| AB | (1,34) | 0.016 | 3.522 | 0.071 |
| BB | (1,34) | 0.134 | 30.463 | <0.001 |
| G: F | (1,34) | 0.036 | 8.204 | 0.008 |
| SM | (1,34) | 0.013 | 2.986 | 0.095 |
| BD | (1,34) | 0.021 | 4.869 | 0.035 |
| pH | (1,34) | 0.003 | 0.570 | 0.456 |
Note: AB, above-ground plant biomass; BB, below-ground plant biomass; G: F, grass: forb biomass ratio; SM, soil moisture; BD, soil bulk density. Bold values represent significant effects (P < 0.05).
4. Discussion
Our study provides clear evidence that the effects of moderate herbivore grazing on the shallow layer of SOC storage strongly depend on herbivore assemblage and grassland type (figures 2 and 6). Although grazing intensity was comparable, the grazing activity of specific herbivore species and assemblages showed distinct effects on storage in various grassland types (figure 2). Specifically, CG increased SOC storage in mesic grasslands, and conversely, SG in the xeric grassland increased SOC storage. MG of sheep and cattle exhibited positive effects on SOC storage in all grasslands.
The differential and idiosyncratic herbivore effects on soil C might be associated with the following three mechanisms: (a) contrasting foraging modes of sheep and cattle resulted in different root growth; (b) contrasting diet selections for sheep and cattle changed differently plant community composition; (c) differences in the trampling effects of sheep and cattle related to herbivore assemblage led to different soil properties. Our results revealed that grazing-induced variations in below-ground root biomass explained most SOC storage variability (figure 6). Substantial and empirical evidence also demonstrated that root litter inputs were conserved in SOC more effectively than shoot litter, as has been reported previously by some authors (e.g. Rasse et al 2005, Wilson et al 2018).
As grazing-induced changes in C inputs are more likely to affect shallow soil layers where the majority of root production occurs, long-term C dynamics within this layer may influence SOC sequestration (McSherry and Ritchie 2013). Our results demonstrate that grazing-induced changes in plant root biomass varied with different grazing assemblages and grassland types. CG greatly increased below-ground biomass in meadow steppe and typical steppe (relatively mesic grasslands) but not in desert steppe (figure 3), which resulted in more C input into soil in relative mesic grasslands (figure 2). Unlike CG, SG significantly increased below-ground biomass in desert steppe (figure 3), thereby improving soil C input (figure 2). Further analysis showed a strong positive relationship between SOC and below-ground biomass (figure 7). We therefore speculate that different foraging strategies of herbivores may cause varying effects of plant compensatory growth and simultaneously the knock-on effects stimulate a shallow layer of root growth. Sheep typically adopt a 'top removal, stem-pulling, and defoliation' foraging strategy, while cattle employ a 'tongue sweeping and teeth cutting' foraging strategy (Feng 2014), which may lead to disparate effects on root growth. In mesic grasslands, where there was a large physical appearance of above-ground plant biomass and a high physical appearance of plant height, not surprisingly, the foraging strategy of cattle enabled the consumption of more above-ground plant biomass. Then, in order to restore growth, the plants may allocate additional photosynthetic products to their roots to obtain more nutrients and water, which consequently induces root growth (Olsen et al 2011). Comparatively, in xeric grasslands, there was a sparse above-ground plant biomass and a dwarf appearance of plant height, and forbs with low biomass accounted for a larger proportion than mesic grasslands. Sheep stimulate more shallow layer root growth than cattle, depending on sheep foraging behavior (Wang et al 2019b). Consequently, sheep and cattle altered C sequestration to varying degrees by stimulating root growth according to grassland type.
Of particular interest here, MG of cattle and sheep greatly increased the below-ground biomass of the shallow layer, and thus increased soil C storage across all grassland types (figure 3). Similarly, the predominantly positive effects of MG on shallow layer soil C storage have been found for the first time in meadow steppe with the high plant diversity community of northern China (Chang et al 2018). Our study further demonstrates this effect in the three main types of eastern Eurasian steppe, including meadow steppe, typical steppe, and desert steppe. In our prior studies, Wang et al (2019a) demonstrated that mixed herbivore species grazing substantially increased ecosystem multifunctionality (including plant biomass production, plant leaf N and P, above-ground insect abundance, nutrient cycling, soil C stocks, water regulation, and plant–microbe symbiosis) by promoting biodiversity (the diversity of plants, insects, soil microbes, and nematodes). Furthermore, an experimental study indicated that MG significantly enhanced plant diversity, which was not observed with isolated cattle or SG in meadow steppe with the high plant diversity community (Liu et al 2015). Afterward, a similar experimental study confirmed that mixed herbivores grazing had a greater effect on soil N availability than single species grazing in meadow steppe with the low plant diversity community (Liu et al 2017). The above works provide considerable experimental evidence that MG may serve as an optimal grazing regime to increase the multifunctionality of grasslands. The advantage of MG is that different herbivore types often have complementary and additive effects, and the ecological relationships between herbivores can be facilitated (Bakker et al 2006). Moreover, the differences in selective foraging between various herbivores might be a mechanism that drives niche separation (consider separation of diets within the same landscape) because patches that are not utilized by large herbivores, due to insufficient size or resource density, may contain several smaller patches with sufficient resource density to be selected by smaller herbivores (Laca et al 2010). The strong stimulation of different herbivore types in idiosyncratic patches is more conducive to root growth, thus increasing soil C storage.
There is a line of evidence that plant community composition affects soil C sequestration (Fornara and Tilman 2008, Cong et al 2014), whereby forbs play a crucial role in C inputs of grazing systems (Bagchi and Ritchie 2010). Our findings demonstrate that different herbivore assemblages had disparate effects on plant functional type (i.e. grass: forb ratio) as the result of foraging behavior, which further affect soil C (Albon et al 2007, Chang et al 2018). Sheep tend to forage on forbs, giving rise to an increase in the grass: forb biomass ratio (Song et al 2017). Chang et al (2018) have also shown that SG reduced forb biomass, this effect occurred only in the high plant diversity community, but did not hold true in the low plant diversity community (Chang et al 2018). In the present study, the results demonstrated that sheep had a significant effect on the grass: forb ratio in mesic grasslands. This may be related to the physiological characteristics of plants under different climatic conditions (Wright et al 2004). In mesic grasslands, forbs tend to have large leaf areas with high nutrient concentrations. Sheep select preferred components from different plant types, therefore foraging on more nutritional portions of plants, which increases the grass: forb ratio (Liu et al 2017). In xeric grassland, cattle prefer to forage on grass to satisfy their own needs, thus reducing the grass: forb ratio (figure 3). This provides a possible explanation as to why cattle have a significant effect on C accumulation in mesic grasslands, while sheep have a potent effect on C accumulation in xeric grassland.
The effects of grazing on soil were also herbivore specific. Trampling by different herbivore assemblages can also have significant and distinct impacts on the shallow layer of soil C sequestration (Vallentine 2000, He et al 2011, Eldridge et al 2016). Several studies compared the trampling effects of sheep and cattle, which revealed the more dramatic effect of cattle trampling on soil compaction compared with sheep. This is likely and obviously due to the much greater cattle body mass and pressure per unit area of hoof intensity than that of sheep under the same grazing pressure (Cumming and Cumming 2003, Eldridge et al 2016). Nonetheless, contradictory to previous studies, we found that sheep trampling intensity was slightly higher than that of cattle, which at first seems counterintuitive (figure 4). This was likely due to the fact that previous studies did not consider the browsing habits of herbivores. Compared with the browsing and trampling effects of cattle, sheep are relatively small in size and highly mobile. Furthermore, sheep have highly selective foraging requirements for plant species, and as a consequence they travel more extensively with more steps. On the contrary, it is not convenient for cattle to travel due to their large body mass. The physiological costs of frequently turning and browsing likely outweigh the perceived benefits for cattle, hence, cattle browsing is restricted to fewer areas and habitats than sheep (Laca et al 2010).
Furthermore, a global meta-analysis suggested that the effects of grazing on soil C sequestration may be contingent on soil texture (McSherry and Ritchie 2013). In mesic grasslands, soil texture tends to be fine textured. Sheep traverse many more steps than do cattle (as explained above), and sheep trampling is not conducive to the shallow layer of SOC sequestration. In contrast, the soil tends to be coarse textured in xeric grasslands, and the damage of cattle trampling is greater than that of sheep, which results in a relatively negative effect on the shallow layer of SOC storage by CG. Overall, the differences in trampling pressure and browsing habits between sheep and cattle have variable effects on soil properties, and subsequently on soil C sequestration. Therefore, it is necessary to further disentangle the comprehensive effects of herbivore assemblages on soil properties to better inform how diverse herbivore assemblages impact C sequestration in different grassland types.
5. Conclusions
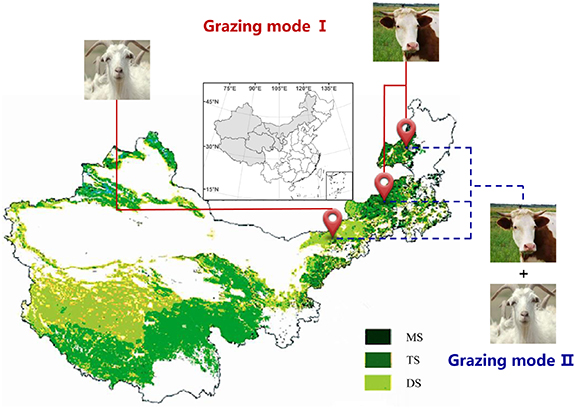
Our findings support the herbivore-size-dependent hypothesis, with herbivore impacts on soil C sequestration in grasslands being dependent upon herbivore type and habitat. Specifically, our study demonstrates that herbivore species-specific grazing can help to improve the shallow layer of soil C sequestration function in grassland ecosystems. Large herbivores and small herbivores are suitable for grazing in mesic grasslands and xeric grasslands, respectively. Moreover, MG may be optimal moderate grazing stewardship across grassland types in northern China (figure 8). Diverse herbivores may be what historically existed in some landscapes, and therefore restoring this type of grazing may be important for restoring ecosystem function and C sequestration. Our study has potentially significant implications for the more effective grazing management of C functionality and services. It suggests that mixed herbivore assemblage should be further considered to comprehensively elucidate and manage the impacts of grazing systems on the C functionality in grassland ecosystems. Furthermore, due to the potential importance of grazing history and subsoil C for the objective evaluation of SOC variability toward the reduction of greenhouse gases, the roles of grazing history and subsoil C would seem to be a fruitful area for future research.
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Figure 8. Promotion of grazing modes for maximum accumulation of SOC in MS, TS, and DS. Herbivore species linked to specific sites represent the recommendation for grazing herbivores in different grassland types.
Download figure:
Standard image High-resolution image{kind=link}
Acknowledgments
We thank D Pan, C He, S Zheng, M, Xu, R Ning, S Jiang, X Yue, and Y Li for help with the fieldwork and laboratory analysis. This project was supported by the National Natural Science Foundation of China (Nos. 31772652, 32061143027), the Program for Introducing Talents to Universities (B16011), the Ministry of Education Innovation Team Development Plan (2013-373), and China Scholarship Council (Grant No. 201806620032).
Data availability statement
The data that support the findings of this study are available upon reasonable request from the authors.
Authors' contributions
JL and HC performed the experiments and led in data analysis and writing efforts; LW and DW designed the experiments and contributed to data analysis and writing; LW, DW, JL, CL, and CW developed the original idea of the experiments; JW, YL, YL, and HW contributed to revise the manuscript critically; SD, XL, YL, and TL performed the experiments. All the authors contributed critically to the drafts and gave final approval for publication.
Conflict of interest
The authors declare no competing interests.
Ethics statement
All experimental procedures were carried out in accordance with the Regulation on the Administration of Laboratory Animals (2017 Revision, State Council, Beijing, China).