 Carolina Peñaloza1
Carolina Peñaloza1 Agustin Barria1
Agustin Barria1 Athina Papadopoulou2
Athina Papadopoulou2 Chantelle Hooper2Joanne Preston3
Chantelle Hooper2Joanne Preston3 Matthew Green2
Matthew Green2 Luke Helmer3,4,5Jacob Kean-Hammerson4
Luke Helmer3,4,5Jacob Kean-Hammerson4 Jennifer C. Nascimento-Schulze2,6
Jennifer C. Nascimento-Schulze2,6 Diana Minardi2
Diana Minardi2 Manu Kumar Gundappa1
Manu Kumar Gundappa1 Daniel J. Macqueen1John Hamilton7
Daniel J. Macqueen1John Hamilton7 Ross D. Houston8*†
Ross D. Houston8*† Tim P. Bean1*†
Tim P. Bean1*†- 1The Roslin Institute and Royal (Dick) School of Veterinary Studies, University of Edinburgh, Edinburgh, United Kingdom
- 2Centre for Environment, Fisheries and Aquaculture Science (Cefas), Weymouth Laboratory, Weymouth, United Kingdom
- 3Institute of Marine Sciences, University of Portsmouth, Portsmouth, United Kingdom
- 4Blue Marine Foundation, London, United Kingdom
- 5Ocean and Earth Science, University of Southampton, Southampton, United Kingdom
- 6College of Life and Environmental Sciences, University of Exeter, Exeter, United Kingdom
- 7Lochnell Oysters, Oban, United Kingdom
- 8Benchmark Genetics, Penicuik, United Kingdom
The European flat oyster (Ostrea edulis) is a bivalve mollusc that was once widely distributed across Europe and represented an important food resource for humans for centuries. Populations of O. edulis experienced a severe decline across their biogeographic range mainly due to overexploitation and disease outbreaks. To restore the economic and ecological benefits of European flat oyster populations, extensive protection and restoration efforts are in place within Europe. In line with the increasing interest in supporting restoration and oyster farming through the breeding of stocks with enhanced performance, the present study aimed to evaluate the potential of genomic selection for improving growth traits in a European flat oyster population obtained from successive mass-spawning events. Four growth-related traits were evaluated: total weight (TW), shell height (SH), shell width (SW) and shell length (SL). The heritability of the growth traits was in the low-moderate range, with estimates of 0.45, 0.37, 0.22, and 0.32 for TW, SH, SW and SL, respectively. A genome-wide association analysis revealed a largely polygenic architecture for the four growth traits, with two distinct QTLs detected on chromosome 4. To investigate whether genomic selection can be implemented in flat oyster breeding at a reduced cost, the utility of low-density SNP panels was assessed. Genomic prediction accuracies using the full density panel were high (> 0.83 for all traits). The evaluation of the effect of reducing the number of markers used to predict genomic breeding values revealed that similar selection accuracies could be achieved for all traits with 2K SNPs as for a full panel containing 4,577 SNPs. Only slight reductions in accuracies were observed at the lowest SNP density tested (i.e., 100 SNPs), likely due to a high relatedness between individuals being included in the training and validation sets during cross-validation. Overall, our results suggest that the genetic improvement of growth traits in oysters is feasible. Nevertheless, and although low-density SNP panels appear as a promising strategy for applying GS at a reduced cost, additional populations with different degrees of genetic relatedness should be assessed to derive estimates of prediction accuracies to be expected in practical breeding programmes.
Introduction
The European flat oyster (Ostrea edulis) was an abundant native bivalve species and an important fishery resource in much of Europe up to the 19th century (Pogoda, 2019). However, populations of O. edulis experienced a severe decline across their biogeographic range due to an array of factors including overfishing and habitat degradation (Thurstan et al., 2013), the subsequent invasion of non-native species (e.g., slipper limpet, Crepidula fornicata) (Helmer et al., 2019; Preston et al., 2020) and pathogenic diseases (Robert et al., 1991; Sas et al., 2020). The continuous decimation of native populations in the Atlantic and Mediterranean seas led to significant changes in oyster production, which progressively shifted towards farming (Korringa, 1976), and eventually to the cultivation of different species including Crassostrea angulata (Boeliguf, 2000) and the non-indigenous Pacific oyster (Crassostrea gigas) (Walne and Helm, 1979; Grizel and Héral, 1991). The Pacific oyster was introduced into Europe for aquaculture purposes owing to its favourable production traits, such as a faster growth rate and higher resistance to the main diseases affecting C. angulata and O. edulis (Grizel and Héral, 1991; Renault et al., 1995). Worldwide oyster production is now dominated by the Pacific oyster (97.7%), while the production of the European flat oyster remains stably low, constituting just ∼0.2% of global production in 2002 (FAO, 2022). Despite the demand for shellfish continues to increase (Botta et al., 2020), the level of O. edulis production is stagnant. One of the main factors that hinders the growth of the industry is the lack of a substantial and steady supply of oyster seed (i.e., juveniles) (see Colsoul et al. (2021) for a review). Hence, the optimization of oyster larval production in hatcheries and spatting ponds is key for future European flat oyster aquaculture, as well as for restoration projects, which are also expected to rely on sustainable sources of juveniles for restocking (Pogoda et al., 2020). Importantly, the artificial propagation of flat oyster seed will facilitate the application of selective breeding programmes. Although selective breeding programmes are typically used to improve aquaculture production, they could also benefit the ecological restoration of O. edulis. If desirable traits such as disease resistance are found to have a strong genetic component, then increased resistance to life-limiting diseases — such as bonamiosis (Naciri-Graven et al., 1998; Culloty et al., 2004) — could potentially be achieved while maintaining the adaptive potential (i.e., genetic diversity) of restored populations.
Selective breeding in oysters has mainly focused on improving meat yield, disease resistance, survival and growth (Toro and Newkirk, 1990; Allen et al., 1993; Ragone Calvo et al., 2003; Ward et al., 2005; Dégremont et al., 2015; De Melo et al., 2016; Proestou et al., 2016; Camara et al., 2017; Zhang et al., 2019), with a recent interest in nutritional content and shell shape (Grizzle et al., 2017; Liu et al., 2019; Meng et al., 2019; Wan et al., 2020; He et al., 2022). Among these traits, growth is comparatively simple to assess and consequently select for using phenotypic information. Although the direct comparison of heritability estimates from different studies is difficult (e.g., due to intrinsic differences between populations), estimates for growth rate in oysters tend to be moderate (e.g., 0.31 and 0.55—Evans and Langdon (2006) and De Melo et al. (2016), respectively). As a result, fast-growing lines of oysters have been developed for some of the main commercial species, such as the Pacific (C. gigas) (Zhang et al., 2019), Portuguese (C. angulata) (Vu et al., 2020), Eastern (C. virginica) (Varney and Wilbur, 2020) and Sydney rock (Saccostrea glomerata) (Fitzer et al., 2019) oyster. Initial attempts to genetically improve the European flat oyster O. edulis resulted in an average 23% increase in growth rate compared to an unselected (control) line (Newkirk and Haley, 1982). This striking genetic response was not replicated in a second generation of selection, possibly due to unintentional inbreeding (Newkirk and Haley, 1983). Indeed, even relatively modest levels of inbreeding have been shown to significantly affect performance traits in oysters (Evans et al., 2004), highlighting the importance of an adequate management of genetic diversity in hatchery-derived stocks. Moreover, oysters and bivalves in general, appear to have a high genetic load [see for a review Plough (2016)] and, therefore, may be particularly susceptible to inbreeding depression. Hence, the incorporation of genomic tools into shellfish breeding schemes will be key for balancing genetic gain with population diversity in order to sustain the long-term progress for traits under selection.
A vast array of genomic tools and resources have become available for genetic research and breeding applications in oysters. For example, for economically relevant species, chromosome-level genome assemblies (Li et al., 2021; Modak et al., 2021; Peñaloza et al., 2021; Qi et al., 2021), SNP arrays (Lapegue et al., 2014; Gutierrez et al., 2017; Qi et al., 2017) and medium-density linkage maps (Jones et al., 2013; Wang et al., 2016; Gutierrez et al., 2020; Li et al., 2018; Yin et al., 2020) have been produced. These resources have been applied to examine the genetic basis of growth (Jones et al., 2014; Gutierrez et al., 2018; He et al., 2021), low salinity tolerance (McCarty et al., 2022), disease resistance (Gutierrez et al., 2020; Yang et al., 2022) and nutritional content (Meng et al., 2019). For the European flat oyster, high-quality genomes have recently been released (Boutet et al., 2022; Gundappa et al., 2022), which along with available high-throughput genotyping techniques (e.g., SNP arrays and genotype-by-sequencing approaches), provide the opportunity for gaining insight into the genomic architecture of relevant production traits. Most of the traits of economic importance in aquaculture species have a polygenic architecture (Zenger et al., 2019). For polygenic traits (i.e., those controlled by many loci), the application of predictive techniques such as genomic selection (GS) may enable a faster genetic gain than conventional pedigree-based selection. GS is a method based on genome-wide markers in which the effect of all loci are simultaneously used for predicting the estimated breeding values (EBV) of selection candidates (Meuwissen et al., 2001), and has shown major potential in aquaculture species, where it can be used to characterise variation within and between large families of potential breeders. However, commercial application to aquaculture production is largely limited to the major finfish and crustacean species (e.g., salmonids, Nile tilapia, shrimp) (Zenger et al., 2019; Lillehammer et al., 2020; Boudry et al., 2021). Studies into the feasibility of applying genomic selection schemes in oyster breeding programmes have shown that for growth (Gutierrez et al., 2018; Vu et al., 2021b), edibility (Vu et al., 2021a), and disease resistance traits (Gutierrez et al., 2020; Vu et al., 2021a), greater genetic gains could be achieved through GS compared to traditional breeding. Nevertheless, the practical application of GS as a selection strategy will likely depend on how cost-effective it is compared to pedigree-based methods. The development of feasible alternatives for reducing genotyping costs, such as using affordable low-density genotyping tools that yield similar accuracies than higher-density panels, will be critical for the potential of GS to be realized by oyster breeding programmes.
In line with the increasing interest in supporting oyster culture and restoration through the breeding of stocks with enhanced performance, the overall aim of this study was to evaluate the potential of GS for the genetic improvement of growth and growth-related (morphometric) traits in the European flat oyster. First, the heritability of total weight, shell length, shell width and shell height was estimated for a hatchery-derived population genotyped using a ∼15K SNP array. Second, a genome-wide association (GWAS) analysis was conducted to dissect the genetic architecture of the measured traits. Last, to evaluate whether GS may be an effective and cost-effective strategy for improving traits associated with oyster growth, the accuracy of genomic predictions using reduced density SNP marker panels was assessed.
Materials and Methods
Field Experiment
The European flat oyster population used in this study was generated in a UK-based hatchery (Seasalter Morecombe hatchery) by mass spawning of approximately 40 broodstock parents over several spawning events. The resulting F1 generation was then deployed to Lochnell oysters (56.494°N, 5.459°W) and grown for 6 months in ortac grow-out cages. Next, animals were transferred to the Institute of Marine Sciences at the University of Portsmouth (UK), and maintained in a flow-through system until deployment. During this holding period, ∼1,000 randomly selected oysters were individually tagged and their first phenotype measurements recorded (see “Phenotypes” section below). Prior to deployment, animals were cleaned of fouling, washed in fresh water and dried. Embossed plastic tags with unique identifier codes were attached with epoxy resin glue. Animals were returned to aquaria within the hour. Oysters were placed in Aquamesh® cages (L 0.55 m × W 0.55 m × D 0.4 m) at a density of 200 oysters per cage, and deployed 1 m below floating pontoons at Port Hamble Marina (MDL) in the River Hamble (50.861°N, 1.312°W) in January 2019. Mortalities were recorded monthly and dead oysters—i.e., those with empty or gaping shells—were removed from the experiment. General disease status was assessed on subsets of oysters throughout the experiment by histology and in situ hybridisation using an adaptation of available methods (Montagnani et al., 2001; Fabioux et al., 2004). In addition, the presence of Bonamia ostreae, a protozoan parasite that causes a lethal infection of flat oyster haemocytes (Pichot et al., 1979), was assessed by qPCR following Robert et al. (2009). The prevalence of B. ostreae infections was negligible; hence disease status had a minor influence on the assessment of growth traits in the experimental population. After 10 months of growth under field conditions, gill tissue was dissected from individuals alive at the end of the study and preserved in molecular grade absolute ethanol (Fisher Scientific) for genetic analysis.
Phenotypes
Four growth-associated traits were measured at three time points over the course of 10 months: total weight (TW, the weight of an individual oyster including the shell), shell length (SL, the maximum distance between the anterior and posterior margins), shell height (SH, the maximum distance between the hinge to the furthermost edge), and shell width (SW, the maximum distance at the thickest part of the two shell valves) (Figure 1). Weight was recorded in grams up to one decimal place. Shell measurements were taken with traceable digital callipers (Fisher Scientific) with 0.02 mm precision. Oysters were cleaned and defouled before measurements were taken.
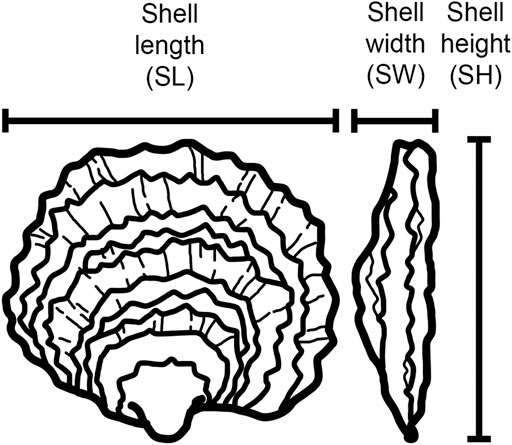
FIGURE 1. Nomenclature of the growth-related morphometric traits measured in this study.
DNA Extraction
Total DNA was isolated from gill tissue following a CTAB (cetyltrimethylammonium bromide)-based extraction protocol [details in Gutierrez et al. (2017)]. The integrity of the extracted DNA was assessed by agarose gel electrophoresis, while DNA purity was verified on a Nanodrop ND-1000 (Thermo Fisher Scientific) spectrophotometer by checking the 260/280 and 260/230 ratios. All samples had 260/280 and 260/230 values ≥ 1.85 and ≥1.96, respectively.
SNP Genotyping and Quality Control
Whole-genome genotyping of ∼15K SNPs was carried out by IdentiGEN (Dublin, Ireland) using the combined-species Affymetrix Axiom oyster SNP-array (Gutierrez et al., 2017). Signal intensity files were imported to the Axiom analysis Suite v4.0.3.3 software for quality control (QC) assessment and genotype calling. Genotypes were generated using the default parameter settings for diploid species, resulting in 11,808 SNPs typed for 870 individuals. To assess the reproducibility of genotype calls, five DNA samples from the same individual were genotyped independently on three different arrays, and their genotype concordance evaluated through an identity-by-state (IBS) analysis. The genotype concordance rate among replicates was 99.7%, demonstrating a high reproducibility of the genotyping assays. The flanking region of these markers were mapped to the O. edulis chromosome-level genome assembly (Gundappa et al., 2022). Of the 11,808 SNPs, 10,025 had uniquely mapping probes and were retained for downstream analysis. A total of 1,539 markers (15.4%) were monomorphic in the population under study. Additional QC was performed on markers and samples using Plink v2.0 (Chang et al., 2015). SNP variants were retained for further analysis if they had a call rate > 95% and a minor allele frequency (MAF) > 0.05. These filters removed 4,391 SNPs (leaving a total of 5,634 SNPs), of which the majority were filtered out based on the MAF threshold (i.e., were monomorphic or near-monomorphic in this population). Given that significant sub-clustering was detected in the data (Supplementary Figure S1), possibly due to a high variance in the reproductive success of broodstock parents and/or temporal variation in spawning, a k-means clustering method was used to assign individuals into groups. Deviations from Hardy-Weinberg Equilibrium (HWE) were tested separately in each of the three genetic clusters identified by the analysis. SNP markers showing significant deviations (HWE p-value < 1e-10) in two of the three clusters were excluded from the analysis. Sample QC included removing individual oysters with a missingness above 5% and high heterozygosity (i.e., more than three median absolute deviations from median). Finally, a principal component analysis (PCA) was performed using a set of ∼3.5K SNPs for which no pair of markers within a window of 200 kb had a
Genetic Parameter Estimation
Genetic parameters for growth-related traits were estimated by fitting the following univariate linear mixed model in GEMMA v0.95alpha (Zhou and Stephens, 2012):
Where y is the vector of observed phenotypes; μ is the overall mean of the phenotype in the population; b is the vector of fixed effects to be fitted (the first five principal components were included as covariates);
Bivariate animal linear models were implemented to estimate the genetic (co)variance between TW, SL, SH and SW. Each bivariate analysis was fitted with the same top 5 PCs mentioned above. Subsequently, genetic correlations among traits were estimated as the ratio of the covariance of two traits to the square root of the product of the variance for each trait. Phenotypic correlations between traits were calculated using the Pearson correlation coefficient.
Genome-Wide Association Study
To identify SNPs in the flat oyster genome correlated with variation in growth-related traits, a GWAS was performed by implementing the same model described previously in the GEMMA software. SNPs were considered significant at the genome-wide level if their likelihood ratio test P-values surpassed a conservative Bonferroni-corrected significance threshold (α/4,577 = 1.09e-5). To derive a threshold for chromosome-wide (suggestive) significance, α was divided by the average number of SNPs per chromosome (α/457 = 1.09e-4). The single-marker P-values obtained from GEMMA were plotted against their chromosome location using the R package qqman v 0.1.4 (Turner, 2018). To assess the inflation of the association statistics, the genomic control coefficient lambda λGC was calculated following Devlin and Roeder (1999). Candidate genes were searched within 100 kb of the most significant SNP loci using BEDOPS v2.4.26 (Neph et al., 2012).
Genomic Prediction
To evaluate the accuracy of genomic selection, a 5-fold cross validation approach — animals split into training (80%) and validation (20%) sets—was used on a population of 840 oysters genotyped for 4,577 informative SNP markers. To reduce stochastic effects arising from individual sampling, each analysis was repeated 10 times. For each replicate, animals were randomly partitioned into five subsets (each subset contained 168 individuals). TW, SL, SH and SW phenotypes recorded in individuals allocated to one of the subsets (validation set) were masked. The breeding values of the validation set were then predicted based on the information from the remaining four subsets (training sets) using model (1). The model was fitted using the AIREMLF90 module from BLUPF90. The accuracy of genomic predictions was calculated as follows:
where rgEBV,y is the correlation between the predicted and the actual phenotypes of the validation set, while
Evaluation of the Effect of SNP Density on Genomic Predictions
To assess the effect of SNP density on the accuracy of genomic predictions of growth-related traits, SNP panels of varying sizes were randomly sampled from the final pool of QC-filtered array markers (n = 4,577 SNPs). Panels of the following densities were evaluated: 4, 3, 2, 1K, 500, 400, 300, 200 and 100 SNPs. To build the lower-density panels, markers were randomly sampled from the full QC-filtered SNP dataset in proportion to chromosome lengths using the R package CVrepGPAcalc v1.0 (Tsairidou, 2019; Tsairidou et al., 2020). To account for sampling bias, 10 SNP panels were generated for each of the SNP densities. The average genomic prediction accuracies of the different low-density panels were compared against the equivalent accuracy values obtained with the full panel.
Results and Discussion
Growth Traits and Heritability
Improvement of growth rate is typically one of the first traits to be included as a selection target in breeding programmes across many farmed species. In this study, oyster growth rate was assessed in a hatchery-derived oyster population that was translocated to a growing site and monitored for 10 months. The experimental population had a lower genome-wide heterozygosity (Ho = 0.27; He = 0.22) compared to the values reported by Vera et al. (2019) (Ho ≥ 0.31) for a diverse set of flat oyster populations genotyped with the same array. An overall mortality of 14% was observed during the field trial, among which the majority (36%) occurred during a summer month (July). At the end of the experimental period, the O. edulis population had the following growth means and standard deviations: +15.7 g (SD = 5.8), 50.8 mm (SD = 7.3), 12.9 mm (SD = 2.3) and 45.8 mm (SD = 8.9), for TW, SH, SW, SL, respectively (Table 1). The phenotypic correlation was found to be the highest (
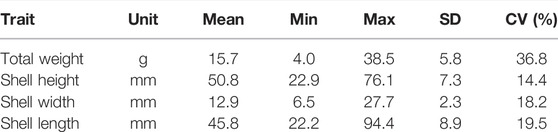
TABLE 1. Summary statistics of the phenotypic data (SD: Standard deviation; CV: coefficient of variation).
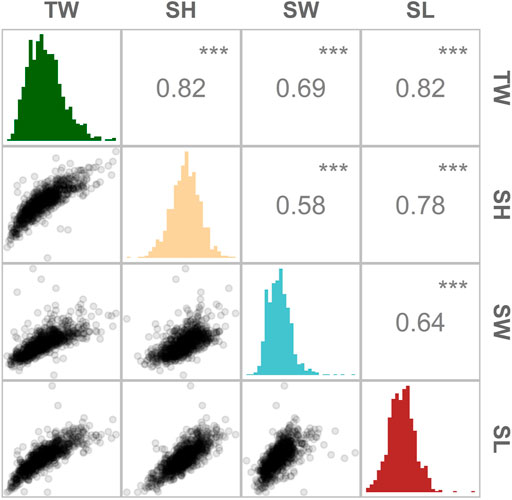
FIGURE 2. Distribution and magnitude of the phenotypic correlations between growth-related traits in Ostrea edulis. Pearson’s correlation between traits (above the diagonal), histogram of trait distribution (diagonal) and scatterplots comparing two traits (below the diagonal). TW (total weight), SL (shell length), SH (shell height) and SW (shell width). *** indicates p-values < 0.001.
For the European oyster population under study, the heritability estimates of these growth-related traits were in the low to moderate range of 0.22 (for SW) to 0.45 (for TW) (Table 2). Consistent with similar studies carried out in related oyster species (Xu et al., 2017; Vu et al., 2020), heritability estimates based on SNP markers were higher for total weight than for growth-related morphometric traits (i.e., shell height, shell width and shell length). The estimation of heritability for total weight (TW) was similar to those reported for nine- month-old Portuguese oysters (

TABLE 2. Estimates of heritability (
Genome-Wide Association Analysis for Growth-Related Traits
A GWAS of ∼4.5K SNPs passing the filtering criteria were genotyped on 840 oysters with phenotypic records to gain insight into the genetic basis of growth rate variation in O. edulis. Three of the four examined traits showed association signals surpassing the genome-wide level of significance (Figure 3). The genomic inflation factor lambda of the GWAS analysis were close to the desired value (λ = 1) (see Supplementary Figure S2), indicating that population structure was adequately accounted for by the model. For TW, the GWAS identified two putative quantitative trait loci (QTLs) on chromosome 4 associated with the trait. The presence of two separate QTLs is supported by the low linkage disequilibrium observed between the most significant SNPs at each locus (pairwise
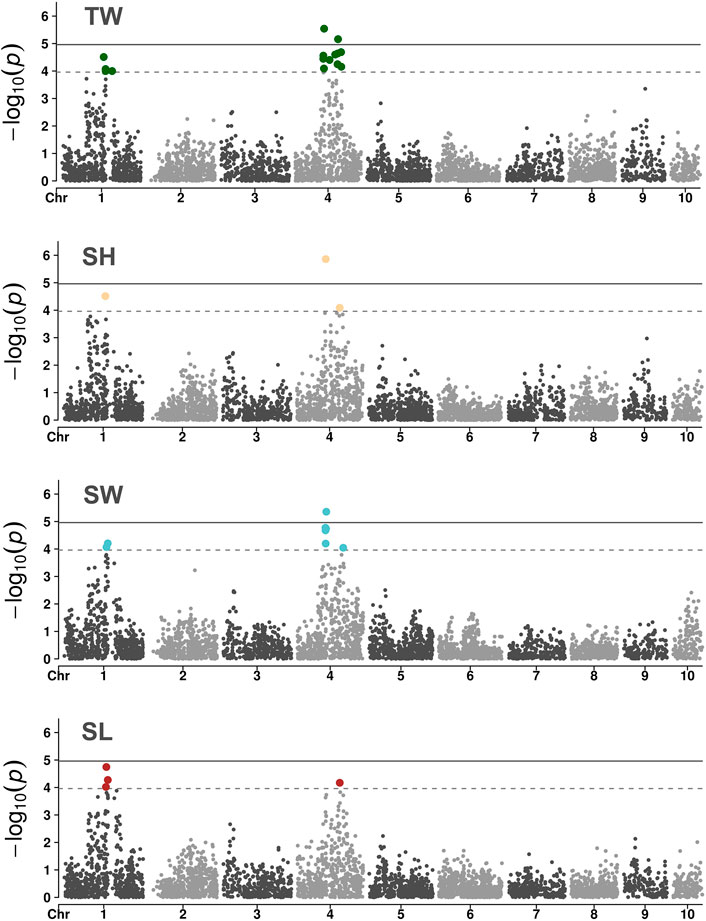
FIGURE 3. Manhattan plots of the GWAS for growth-related traits in a European flat oyster population. Solid lines indicate the threshold value for genome-wide significance. Dashed lines indicate the threshold for a suggestive (chromosome-level) significance.
The marker showing the most significant association with TW, SH and SW is located in the exon of a gene annotated as a N4BP2 (NEDD4 Binding Protein 2)-like protein (Gene ID: FUN_017843; Gundappa et al., 2022). The predicted protein product of this gene contains a highly conserved AAA domain, able to bind and hydrolyse ATP (Lupas and Martin, 2002). Proteins with these domains have been shown to be involved in several cell processes, including protein folding, proteolysis and membrane fusion. Further characterization of this N4BP2-like protein would help better understand the genetic component of growth variation in oysters. Nevertheless, considering that the candidate marker on N4BP2 explained a small percentage of the phenotypic variance, independent oyster populations should first be evaluated to confirm the validity of the association signal. A second genome-wide significant association—detected only in the TW GWAS—was located in the exon of an uncharacterized gene (FUN_018833) whose product displays > 80% amino acid identity and 99% coverage (best BLASTp hit to NCBI's nr database) with similarly uncharacterized proteins in C. gigas and C. virginica (NCBI accession numbers XP_011433755.2 and XP_022325737.1, respectively). Additional genes within the two genomic regions (+/- 100 kb) showing significant associations with flat oyster growth traits are shown in Supplementary Table S2. Given that the SNPs identified in this study had a small effect on the traits in question, GS would be an effective approach for increasing genetic gains from selection.
Genomic Selection
The incorporation of genetic markers into breeding programmes requires a previous understanding of the genetic architecture of the targeted trait(s). In the O. edulis population under study, the genetic contribution to the observed variation in growth-related traits was largely polygenic in nature. For the improvement of polygenic traits, genomic selection has been shown to be superior to alternative marker-aided selection due to genome-wide markers capturing a higher proportion of the genetic variation in a trait compared to individual QTL-targeted markers. Consequently, by means of applying GS, higher predictions have been achieved for several production traits in a wide range of commercially important aquaculture species [reviewed in Houston et al. (2020)]. Despite GS not yet being widely operational in oyster breeding programmes (Boudry et al., 2021), studies have demonstrated the potential of incorporating genome-wide information into selection schemes in these taxa. In the Pacific oyster, Gutierrez et al. (2018) showed that prediction accuracies for growth-traits increased ∼25–30% when the genetic merit of individuals was estimated from SNP markers using the Genomic Best Linear Unbiased Prediction (GBLUP) model (VanRaden, 2008) compared to a classical pedigree-based approach (PBLUP). Similar results were reported in the Portuguese oyster, as prediction accuracies increased 7–42% for growth-related traits when EBVs obtained by GBLUP were compared to those obtained by PBLUP (Vu et al., 2021a). Since the flat oyster population under study derived from a mass-spawning event, the pedigree structure was unknown. Therefore, comparisons between pedigree and genome based methods for estimating breeding values (e.g., GBLUP and Bayesian approaches) could not be performed.
One of the major barriers of implementing GS is the high number of markers required to accurately predict EBVs and the cost of genotyping these markers (Goddard and Hayes, 2007). Therefore, the design of a strategy to reduce the cost of genotyping is critical for the extensive adoption of genomic prediction approaches in aquaculture breeding programmes. One such strategy involves genotyping the minimum number of markers required to achieve maximal accuracy, which by definition is equal to that obtained with a full panel of markers. As shown by Kriaridou et al. (2020) for different aquaculture species, the use of low-density SNP panels has the potential to achieve similar EBV accuracies as when using medium density genotype datasets of around 7–14K SNPs. The authors estimated that only 1,000 to 2,000 SNPs are required to achieve maximal accuracy. These results were shown to be consistent across a range of traits (e.g., disease resistance, growth) and species (e.g., Pacific oyster, Atlantic salmon) showing robustness to differences in family structure, genotyping approach, trait heritability and the underlying genetic architecture. In agreement with these findings, maximal accuracy was attained herein for all the assessed growth-related traits at a minimum density of 2K SNPs, with only a slight decline in accuracy observed at the lower densities evaluated (Figure 4A). Consequently, a reduction in the costs of applying GS for improving growth traits in O. edulis can be achieved by means of exploiting low-density SNP panels. Although low-density panels might not accurately capture the genetic resemblance among individuals within a population, and therefore show reduced genetic variance estimations and EBV predictions when compared with high density panels, their use has been widely evaluated and suggested for different aquaculture species and traits. Furthermore, studies have shown that low-density panels can achieve higher accuracies than the classical pedigree-based approach, being a feasible alternative to identify candidates with the highest genetic merit. For example, in rainbow trout (Oncorhynchus mykiss) Vallejo et al. (2018) showed that at least 200 SNPs could exceed PBLUP accuracies for bacterial cold water disease resistance. Whilst Al-Tobasei et al. (2021) found a similar trend when using between 500–1000 SNPs for fillet yield traits. To date, the utilization of low-density panels to decrease the cost of genomic evaluations has also been tested in several aquaculture species, including Atlantic salmon (Salmo salar) (Tsai et al., 2016; Correa et al., 2017), rainbow trout (Yoshida et al., 2018; Al-Tobasei et al., 2021), and Nile tilapia (Oreochromis niloticus) (Yoshida et al., 2019; Barría et al., 2021), suggesting that the development of cost-effective strategies for applying GS will be key for shaping modern aquaculture breeding programs.

FIGURE 4. Evaluation of the effect of SNP density on genomic predictions of growth related traits in a European flat oyster population. (A) Percentage of the maximum genomic prediction accuracy achieved using different lower density SNP panels. Values were calculated by dividing the mean accuracy (averaged over ten replicates) estimated at each nominal SNP density by the accuracy obtained using the full SNP dataset. (B) Average genomic prediction accuracy values obtained for oyster growth traits at different panel densities.
Although our results highlight the possibility of reducing the genotyping costs associated with genomic prediction approaches, caution should be taken as even for the smallest marker density (i.e., 100 SNPs), prediction accuracies (averaged over ten replicates) were high and close to the value obtained with the full marker panel. By using only 100 SNPs the estimated decrease in the accuracy of genomic breeding values (GEBVs) was of 5% for SL, 7% for TW and SW, and 10% for SH (Figure 4A). These values highly exceed those reported in the literature for aquaculture species, where reductions > 20% were estimated for panels with 100 SNPs compared to a complete dataset (Kriaridou et al., 2020). The relative stability of GEBVs observed across different marker densities (Figure 4B) is likely explained by the underlying genetic structure of the dataset. For this study, 40 potential parents were placed in the same tank and spawned during successive events. The genetic analysis of the progeny revealed that the population was dominated (70% of the sample size; n = 589) by a group of highly related individuals (Supplementary Figures S1, S3), suggesting there was a large variance in reproductive success among breeders, as previously reported in mass spawning of oysters (Lallias et al., 2010). In the context of GS, the inclusion of highly related animals in the training and validation sets results in only a small number of markers being required to capture the haplotype effects, as related animals share longer haplotypes (Hickey et al., 2014). The fact that in the current study animals grouped in the reference and validation data sets were highly related would have likely increased the accuracy of predicted GEBVs, even when animals are genotyped at low density, as also shown by Fraslin et al. (2022) in Atlantic salmon. Therefore, additional populations with different effective population sizes, genetic backgrounds, and degrees of relatedness should be assessed to obtain estimates to be expected in practical breeding programs. Future work focused on evaluating the extent to which low-density panels and alternative strategies (e.g., genotype imputation) can be used to reduce genotyping costs will be key for the cost-effective exploitation of GS by oyster breeding programmes.
Conclusion
Growth-related traits in O. edulis had moderate to low heritability estimates, ranging from 0.22 (for SW) to 0.45 (for TW). High genetic correlations were identified between all traits (> 0.9); hence, TW—a trait easier to measure—can potentially be used as a proxy phenotype for improving the three examined morphometric traits (SH, SW and SL). The GWAS results revealed that growth traits were largely polygenic, but with two distinct QTLs on chromosome 4 reaching genome-wide significance. Prediction accuracies were high for all traits (> 0.83), with minimal differences observed when comparing estimates obtained using different marker densities. Altogether, these results suggest that the high prediction accuracies found in this study could have been influenced by the highly unbalanced family structure of the experimental population. Consequently, although low-density SNP panels appear as a promising cost-effective GS strategy, additional populations with different degrees of genetic relatedness should be assessed to derive estimates of prediction accuracies to be expected in practical breeding programmes in oysters.
Data Availability Statement
The data presented in this study are available in the Mendeley Data repository as independent genotype and phenotype datasets accessible at https://doi.org/10.17632/v7ppb2xr99.2 and https://doi.org/10.17632/sdtjyys7gr.1, respectively.
Author Contributions
TB and RH conceptualized the study. JH provided the animals for the study. CP, CH, DM, TB, LH, JKH, JP and JN performed the experiment including sample collection and recording phenotype information. AP extracted the DNA used for genotyping. CP and AB performed the analysis. MG, DM and CH evaluated disease status. MKG and DJM provided intellectual input to the analysis. CP and AB wrote the manuscript with input from all authors.
Funding
The authors acknowledge funding from the Biotechnology and Biological Sciences Research Council (BBSRC), including Institute Strategic Programme grants (BBS/E/D/20002172, BBS/E/D/30002275 and BBS/E/D/10002070), a grant within the AquaLeap project (BB/S004181/1), funding from Sustainable Aquaculture Innovation Centre (SAIC), Blue Marine Foundation and National Fish and Wildlife Foundation (NFWF).
Conflict of Interest
Author RH was employed by Benchmark Genetics. Author JH was employed by Lochnell oysters.
The remaining authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Publisher’s Note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Acknowledgments
The authors would like to acknowledge MDL Port Hamble Marina for allowing positioning of oyster cages within the Marina, and thank Eric Harris-Scott, Matthew Sanders, Monica Fabra, Tim Regan and Zenaba Khatir for their invaluable help with setup and sampling of the field experiment. For the purpose of open access, the author has applied a CC BY public copyright licence to any Author Accepted Manuscript version arising from this submission.
Supplementary Material
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fgene.2022.926638/full#supplementary-material
Supplementary Figure S1 | PCA of the O.edulis population under study showing the dominance of single cluster comprised of highly related individuals (clst_0).
Supplementary Figure S2 | Quantile-quantile (Q-Q) plots showing the distribution of the expected (red dashed line) versus observed P-values of the GWAS of growth-related traits.
Supplementary Figure S3 | Example of the pairwise kinship coefficient between individuals in the training and validation set across randomly selected iterations of different 5-fold cross validation (CV) replicates (ten replicates in total). Boxplots show the distribution of values for pairs of individuals (one from the training set and one from the validation set) belonging to the same or different population clusters. Relatedness was inferred using the KING-robust method implemented in Plink v2.0.
References
Allen, S. K., Gaffney, P. M., and Ewart, J. W. (1993). Genetic Improvement of the Eastern Oyster for Growth and Disease Resistance in the Northeast. Massachusetts: Northeastern Regional Aquaculture Center, 210.
Al-Tobasei, R., Ali, A., Garcia, A. L. S., Lourenco, D., Leeds, T., and Salem, M. (2021). Genomic Predictions for Fillet Yield and Firmness in Rainbow Trout Using Reduced-Density SNP Panels. BMC Genomics 22, 92. doi:10.1186/s12864-021-07404-9
Barría, A., Benzie, J. A. H., Houston, R. D., De Koning, D.-J., and de Verdal, H. (2021). Genomic Selection and Genome-wide Association Study for Feed-Efficiency Traits in a Farmed Nile Tilapia (Oreochromis niloticus) Population. Front. Genet. 12, 737906. doi:10.3389/fgene.2021.737906
Botta, R., Asche, F., Borsum, J. S., and Camp, E. V. (2020). A Review of Global Oyster Aquaculture Production and Consumption. Mar. Policy 117, 103952. doi:10.1016/j.marpol.2020.103952
Boudry, P., Allal, F., Aslam, M. L., Bargelloni, L., Bean, T. P., Brard-Fudulea, S., et al. (2021). Current Status and Potential of Genomic Selection to Improve Selective Breeding in the Main Aquaculture Species of International Council for the Exploration of the Sea (ICES) Member Countries. Aquac. Rep. 20, 100700. doi:10.1016/j.aqrep.2021.100700
Boutet, I., Monteiro, H. J. A., Takeuchi, T., Baudry, L., Bonnivard, E., Billoud, B., et al. (2022). Chromosomal Assembly of the Flat Oyster (Ostrea Edulis L.) Genome as a New Genetic Resource for Aquaculture. bioRxiv [Preprint]. Available at: https://doi.org/10.1101/2022.06.26.497643. (Accessed June 29, 2022).
Camara, M. D., Yen, S., Kaspar, H. F., Kesarcodi-Watson, A., King, N., Jeffs, A. G., et al. (2017). Assessment of Heat Shock and Laboratory Virus Challenges to Selectively Breed for Ostreid Herpesvirus 1 (OsHV-1) Resistance in the Pacific Oyster, Crassostrea gigas. Aquaculture 469, 50–58. doi:10.1016/j.aquaculture.2016.11.031
Chang, C. C., Chow, C. C., Tellier, L. C., Vattikuti, S., Purcell, S. M., and Lee, J. J. (2015). Second-generation PLINK: Rising to the Challenge of Larger and Richer Datasets. GigaSci 4, 7. doi:10.1186/s13742-015-0047-8
Colsoul, B., Boudry, P., Pérez‐Parallé, M. L., Bratoš Cetinić, A., Hugh‐Jones, T., Arzul, I., et al. (2021). Sustainable Large‐scale Production of European Flat Oyster ( Ostrea edulis) Seed for Ecological Restoration and Aquaculture: A Review. Rev. Aquac. 13, 1423–1468. doi:10.1111/raq.12529
Correa, K., Bangera, R., Figueroa, R., Lhorente, J. P., and Yáñez, J. M. (2017). The Use of Genomic Information Increases the Accuracy of Breeding Value Predictions for Sea Louse (Caligus rogercresseyi) Resistance in Atlantic Salmon (Salmo salar). Genet. Sel. Evol. 49, 15. doi:10.1186/s12711-017-0291-8
Culloty, S. C., Cronin, M. A., and Mulcahy, M. F. (2004). Potential Resistance of a Number of Populations of the Oyster Ostrea edulis to the Parasite Bonamia ostreae. Aquaculture 237, 41–58. doi:10.1016/j.aquaculture.2004.04.007
Dégremont, L., Nourry, M., and Maurouard, E. (2015). Mass Selection for Survival and Resistance to OsHV-1 Infection in Crassostrea gigas Spat in Field Conditions: Response to Selection after Four Generations. Aquaculture 446, 111–121. doi:10.1016/j.aquaculture.2015.04.029
De Melo, C. M. R., Durland, E., and Langdon, C. (2016). Improvements in Desirable Traits of the Pacific Oyster, Crassostrea gigas, as a Result of Five Generations of Selection on the West Coast, USA. Aquaculture 460, 105–115. doi:10.1016/j.aquaculture.2016.04.017
Devlin, B., and Roeder, K. (1999). Genomic Control for Association Studies. Biometrics 55, 997–1004. doi:10.1111/j.0006-341x.1999.00997.x
Evans, S., and Langdon, C. (2006). Direct and indirect responses to selection on individual body weight in the Pacific oyster (Crassostrea gigas). Aquaculture 261, 522–534. doi:10.1016/j.aquaculture.2006.07.037
Evans, F., Matson, S., Brake, J., and Langdon, C. (2004). The Effects of Inbreeding on Performance Traits of Adult Pacific Oysters (Crassostrea gigas). Aquaculture 230, 89–98. doi:10.1016/j.aquaculture.2003.09.023
Fabioux, C., Huvet, A., Lelong, C., Robert, R., Pouvreau, S., Daniel, J. Y., et al. (2004). Oyster Vasa-like Gene as a Marker of the Germline Cell Development in Crassostrea gigas. Biochem. Biophys. Res. Commun. 320, 592–598. doi:10.1016/j.bbrc.2004.06.009
FAO (2022). Cultured Aquatic Species Information Programme (CASIP) Ostrea Edulis Linnaeus 1758. https://www.fao.org/fishery/en/culturedspecies/ostrea_edulis/en. (Accessed June 26, 2022).
Fitzer, S. C., McGill, R. A. R., Torres Gabarda, S., Hughes, B., Dove, M., O'Connor, W., et al. (2019). Selectively Bred Oysters Can Alter Their Biomineralization Pathways, Promoting Resilience to Environmental Acidification. Glob. Change Biol. 25, 4105–4115. doi:10.1111/gcb.14818
Fraslin, C., Yáñez, J. M., Robledo, D., and Houston, R. D. (2022). The Impact of Genetic Relationship between Training and Validation Populations on Genomic Prediction Accuracy in Atlantic Salmon. Aquac. Rep. 23, 101033. doi:10.1016/j.aqrep.2022.101033
Goddard, M. E., and Hayes, B. J. (2007). Genomic Selection. J. Anim. Breed. Genet. 124, 323–330. doi:10.1111/j.1439-0388.2007.00702.x
Grizel, H., and Héral, M. (1991). Introduction into France of the Japanese Oyster (Crassostrea gigas). ICES J. Mar. Sci. 47, 399–403. doi:10.1093/icesjms/47.3.399
Grizzle, R. E., Ward, K. M., Peter, C. R., Cantwell, M., Katz, D., and Sullivan, J. (2017). Growth, Morphometrics and Nutrient Content of Farmed Eastern Oysters, Crassostrea Virginica (Gmelin), in New Hampshire, USA. Aquac. Res. 48, 1525–1537. doi:10.1111/are.12988
Gundappa, M. K., Peñaloza, C., Regan, T., Boutet, I., Tanguy, A., Houston, R. D., et al. (2022). A Chromosome Level Reference Genome for European Flat Oyster (Ostrea Edulis L.) bioRxiv [Preprint]. Available at: https://doi.org/10.1101/2022.06.26.497633. (Accessed June 29, 2022)
Gutierrez, A. P., Turner, F., Gharbi, K., Talbot, R., Lowe, N. R., Peñaloza, C., et al. (2017). Development of a Medium Density Combined-Species SNP Array for Pacific and European Oysters (Crassostrea gigas and Ostrea edulis). G3 (Bethesda) 7, 2209–2218. doi:10.1534/g3.117.041780
Gutierrez, A. P., Matika, O., Bean, T. P., and Houston, R. D. (2018). Genomic Selection for Growth Traits in Pacific Oyster (Crassostrea gigas): Potential of Low-Density Marker Panels for Breeding Value Prediction. Front. Genet. 9, 391. doi:10.3389/fgene.2018.00391
Gutierrez, A. P., Symonds, J., King, N., Steiner, K., Bean, T. P., and Houston, R. D. (2020). Potential of Genomic Selection for Improvement of Resistance to Ostreid Herpesvirus in Pacific Oyster ( Crassostrea gigas ). Anim. Genet. 51, 249–257. doi:10.1111/age.12909
He, X., Li, C., Qi, H., Meng, J., Wang, W., Wu, F., et al. (2021). A Genome-wide Association Study to Identify the Genes Associated with Shell Growth and Shape-Related Traits in Crassostrea gigas. Aquaculture 543, 736926. doi:10.1016/j.aquaculture.2021.736926
He, X., Wu, F., Qi, H., Meng, J., Wang, W., Liu, M., et al. (2022). Whole-genome Resequencing Reveals the Single Nucleotide Polymorphisms Associated with Shell Shape in Crassostrea gigas. Aquaculture 547, 737502. doi:10.1016/j.aquaculture.2021.737502
Helmer, L., Farrell, P., Hendy, I., Harding, S., Robertson, M., and Preston, J. (2019). Active Management is Required to Turn the Tide for Depleted Ostrea edulis Stocks from the Effects of Overfishing, Disease and Invasive Species. PeerJ 7, e6431. doi:10.7717/peerj.6431
Hickey, J. M., Dreisigacker, S., Crossa, J., Hearne, S., Babu, R., Prasanna, B. M., et al. (2014). Evaluation of Genomic Selection Training Population Designs and Genotyping Strategies in Plant Breeding Programs Using Simulation. Crop Sci. 54, 1476–1488. doi:10.2135/cropsci2013.03.0195
Houston, R. D., Bean, T. P., Macqueen, D. J., Gundappa, M. K., Jin, Y. H., Jenkins, T. L., et al. (2020). Harnessing Genomics to Fast-Track Genetic Improvement in Aquaculture. Nat. Rev. Genet. 21, 389–409. doi:10.1038/s41576-020-0227-y
Jones, D. B., Jerry, D. R., Khatkar, M. S., Raadsma, H. W., and Zenger, K. R. (2013). A High-Density SNP Genetic Linkage Map for the Silver-Lipped Pearl Oyster, Pinctada maxima: A Valuable Resource for Gene Localisation and Marker-Assisted Selection. BMC Genomics 14, 810. doi:10.1186/1471-2164-14-810
Jones, D. B., Jerry, D. R., Khatkar, M. S., Moser, G., Raadsma, H. W., Taylor, J. J., et al. (2014). Determining Genetic Contributions to Host Oyster Shell Growth: Quantitative Trait Loci and Genetic Association Analysis for the Silver-Lipped Pearl Oyster, Pinctada maxima. Aquaculture 434, 367–375. doi:10.1016/j.aquaculture.2014.08.040
Kriaridou, C., Tsairidou, S., Houston, R. D., and Robledo, D. (2020). Genomic Prediction Using Low Density Marker Panels in Aquaculture: Performance across Species, Traits, and Genotyping Platforms. Front. Genet. 11, 124. doi:10.3389/fgene.2020.00124
Kube, P., Cunningham, M., Dominik, S., Parkinson, S., Finn, B., Henshall, J., et al. (2011). Enhancement of the Pacific Oyster Selective Breeding Program. Australia: FRDC and Seafood CRC.
Lallias, D., Taris, N., Boudry, P., Bonhomme, F., and Lapègue, S. (2010). Variance in the Reproductive Success of Flat Oyster Ostrea edulis L. Assessed by Parentage Analyses in Natural and Experimental Conditions. Genet. Res. 92, 175–187. doi:10.1017/S0016672310000248
Lapegue, S., Harrang, E., Heurtebise, S., Flahauw, E., Donnadieu, C., Gayral, P., et al. (2014). Development of SNP-Genotyping Arrays in Two Shellfish Species. Mol. Ecol. Resour. 14, 820–830. doi:10.1111/1755-0998.12230
Li, C., Wang, J., Song, K., Meng, J., Xu, F., Li, L., et al. (2018). Construction of a High-Density Genetic Map and Fine QTL Mapping for Growth and Nutritional Traits of Crassostrea gigas. BMC Genomics 19, 626. doi:10.1186/s12864-018-4996-z
Li, A., Dai, H., Guo, X., Zhang, Z., Zhang, K., Wang, C., et al. (2021). Genome of the Estuarine Oyster Provides Insights into Climate Impact and Adaptive Plasticity. Commun. Biol. 4, 1287. doi:10.1038/s42003-021-02823-6
Lillehammer, M., Bangera, R., Salazar, M., Vela, S., Erazo, E. C., Suarez, A., et al. (2020). Genomic Selection for White Spot Syndrome Virus Resistance in Whiteleg Shrimp Boosts Survival under an Experimental Challenge Test. Sci. Rep. 10, 20571. doi:10.1038/s41598-020-77580-3
Liu, S., Li, L., Zhang, S., Wang, W., Yang, J., and Zhang, G. (2019). Heritability Estimates for Nutritional Quality‐related Traits of the Pacific Oyster, Crassostrea gigas. J. World Aquacult. Soc. 50, 738–748. doi:10.1111/jwas.12588
Lupas, A., and Martin, J. (2002). AAA Proteins. Curr. Opin. Struct. Biol. 12, 746–753. doi:10.1016/s0959-440x(02)00388-3
McCarty, A. J., Allen, S. K., and Plough, L. V. (2022). Genome-wide Analysis of Acute Low Salinity Tolerance in the Eastern Oyster Crassostrea virginica and Potential of Genomic Selection for Trait Improvement. G3 (Bethesda) 12, jkab368. doi:10.1093/g3journal/jkab368
Meng, J., Song, K., Li, C., Liu, S., Shi, R., Li, B., et al. (2019). Genome-wide Association Analysis of Nutrient Traits in the Oyster Crassostrea gigas: Genetic Effect and Interaction Network. BMC Genomics 20, 625. doi:10.1186/s12864-019-5971-z
Meuwissen, T. H. E., Hayes, B. J., and Goddard, M. E. (2001). Prediction of Total Genetic Value Using Genome-wide Dense Marker Maps. Genetics 157, 1819–1829. doi:10.1093/genetics/157.4.1819
Mizuta, D. D., and Wikfors, G. H. (2019). Seeking the Perfect Oyster Shell: A Brief Review of Current Knowledge. Rev. Aquacult. 11, 586–602. doi:10.1111/raq.12247
Modak, T. H., Literman, R., Puritz, J. B., Johnson, K. M., Roberts, E. M., Proestou, D., et al. (2021). Extensive Genome-wide Duplications in the Eastern Oyster (Crassostrea virginica). Phil. Trans. R. Soc. B 376, 20200164. doi:10.1098/rstb.2020.0164
Montagnani, C., Le Roux, F., Berthe, F., and Escoubas, J.-M. (2001). Cg-TIMP, an Inducible Tissue Inhibitor of Metalloproteinase from the Pacific oyster Crassostrea gigas with a Potential Role in Wound Healing and Defense Mechanisms1. FEBS Lett. 500, 64–70. doi:10.1016/s0014-5793(01)02559-5
Naciri-Graven, Y., Martin, A.-G., Baud, J.-P., Renault, T., and Gérard, A. (1998). Selecting the Flat Oyster Ostrea edulis (L.) for Survival when Infected with the Parasite Bonamia ostreae. J. Exp. Mar. Biol. Ecol. 224, 91–107. doi:10.1016/S0022-0981(97)00171-8
Neph, S., Kuehn, M. S., Reynolds, A. P., Haugen, E., Thurman, R. E., Johnson, A. K., et al. (2012). BEDOPS: High-Performance Genomic Feature Operations. Bioinformatics 28, 1919–1920. doi:10.1093/bioinformatics/bts277
Newkirk, G. F., and Haley, L. E. (1982). Progess in Selection for Growth Rate in the European Oyster Ostrea edulis. Mar. Ecol. Prog. Ser. 10, 3. doi:10.3354/meps010077
Newkirk, G. F., and Haley, L. E. (1983). Selection for Growth Rate in the European Oyster, Ostrea edulis: Response of Second Generation Groups. Aquaculture 33, 149–155. doi:10.1016/0044-8486(83)90396-4
Peñaloza, C., Gutierrez, A. P., Eöry, L., Wang, S., Guo, X., Archibald, A. L., et al. (2021). A Chromosome-Level Genome Assembly for the Pacific Oyster Crassostrea gigas. GigaScience 10, giab020. doi:10.1093/gigascience/giab020
Pichot, Y., Comps, M., Tige, G., Grizel, H., and Rabouin, M. A. (1979). Recherches sur Bonamia ostreae gen. n., sp. n., parasite nouveau de l' huitre plate Ostrea edulis L. [France]. Rev. Trav. Inst. Peches Marit. 43, 131–140.
Plough, L. V. (2016). Genetic Load in Marine Animals: a Review. Curr. Zool. 62, 567–579. doi:10.1093/cz/zow096
Pogoda, B., Boudry, P., Bromley, C., Cameron, T. C., Colsoul, B., Donnan, D., et al. (2020). NORA Moving Forward: Developing an Oyster Restoration Network in Europe to Support the Berlin Oyster Recommendation. Aquat. Conserv. Mar. Freshw. Ecosyst. 30, 2031–2037. doi:10.1002/aqc.3447
Pogoda, B. (2019). Current Status of European Oyster Decline and Restoration in Germany. Humanities 8, 9. doi:10.3390/h8010009
Preston, J., Fabra, M., Helmer, L., Johnson, E., Harris‐Scott, E., and Hendy, I. W. (2020). Interactions of Larval Dynamics and Substrate Preference Have Ecological Significance for Benthic Biodiversity and Ostrea edulis Linnaeus, 1758 in the Presence of Crepidula Fornicata. Aquat. Conserv. Mar. Freshw. Ecosyst. 30, 2133–2149. doi:10.1002/aqc.3446
Proestou, D. A., Vinyard, B. T., Corbett, R. J., Piesz, J., Allen, S. K., Small, J. M., et al. (2016). Performance of Selectively-Bred Lines of Eastern Oyster, Crassostrea virginica, across Eastern US Estuaries. Aquaculture 464, 17–27. doi:10.1016/j.aquaculture.2016.06.012
Qi, H., Song, K., Li, C., Wang, W., Li, B., Li, L., et al. (2017). Construction and Evaluation of a High-Density SNP Array for the Pacific Oyster (Crassostrea gigas). PLoS One 12, e0174007. doi:10.1371/journal.pone.0174007
Qi, H., Li, L., and Zhang, G. (2021). Construction of a Chromosome‐level Genome and Variation Map for the Pacific Oyster Crassostrea gigas. Mol. Ecol. Resour. 21, 1670–1685. doi:10.1111/1755-0998.13368
Ragone Calvo, L. M., Calvo, G. W., and Burreson, E. M. (2003). Dual Disease Resistance in a Selectively Bred Eastern Oyster, Crassostrea virginica, Strain Tested in Chesapeake Bay. Aquaculture 220, 69–87. doi:10.1016/S0044-8486(02)00399-X
Renault, T., Cochennec, N., and Grizel, H. (1995). Bonamia Ostreae, Parasite of the European Flat Oyster, Ostrea edulis, Does Not Experimentally Infect the Japanese Oyster, Crassostrea Gigas. Bull. Eur. Ass. Fish. Pathol. 15, 78.
Robert, R., Borel, M., Pichot, Y., and Trut, G. (1991). Growth and Mortality of the European oyster Ostrea edulis in the Bay of Arcachon (France). Aquat. Living Resour. 4, 265–274. doi:10.1051/alr:1991028
Robert, M., Garcia, C., Chollet, B., Lopez-Flores, I., Ferrand, S., François, C., et al. (2009). Molecular Detection and Quantification of the Protozoan Bonamia ostreae in the Flat Oyster, Ostrea edulis. Mol. Cell. Probes 23, 264–271. doi:10.1016/j.mcp.2009.06.002
Sas, H., Deden, B., Kamermans, P., zu Ermgassen, P. S. E., Pogoda, B., Preston, J., et al. (2020). Bonamia Infection in Native Oysters (Ostrea edulis) in Relation to European Restoration Projects. Aquat. Conserv. Mar. Freshw. Ecosyst. 30, 2150–2162. doi:10.1002/aqc.3430
Thurstan, R. H., Hawkins, J. P., Raby, L., and Roberts, C. M. (2013). Oyster (Ostrea edulis) Extirpation and Ecosystem Transformation in the Firth of Forth, Scotland. J. Nat. Conserv. 21, 253–261. doi:10.1016/j.jnc.2013.01.004
Toro, J., and Newkirk, G. (1990). Divergent Selection for Growth Rate in the European Oyster Ostrea edulis Response to Selection and Estimation of Genetic Parameters. Mar. Ecol. Prog. Ser. 62, 219–227. doi:10.3354/meps062219
Tsai, H.-Y., Hamilton, A., Tinch, A. E., Guy, D. R., Bron, J. E., Taggart, J. B., et al. (2016). Genomic Prediction of Host Resistance to Sea Lice in Farmed Atlantic Salmon Populations. Genet. Sel. Evol. 48, 47. doi:10.1186/s12711-016-0226-9
Tsairidou, S., Hamilton, A., Robledo, D., Bron, J. E., and Houston, R. D. (2020). Optimizing Low-Cost Genotyping and Imputation Strategies for Genomic Selection in Atlantic Salmon. G3 (Bethesda) 10, 581–590. doi:10.1534/g3.119.400800
Tsairidou, S. (2019). CVrepGPAcalc. Available at: https://github.com/SmaragdaT/CVrep/tree/master/CVrepGPAcalc (Accessed March 15, 2020).
Turner, S. D. (2018). Qqman: An R Package for Visualizing GWAS Results Using Q-Q and Manhattan Plots. J. Open Source Softw. 3 (25), 731. doi:10.21105/joss.00731
Vallejo, R. L., Silva, R. M. O., Evenhuis, J. P., Gao, G., Liu, S., Parsons, J. E., et al. (2018). Accurate Genomic Predictions for BCWD Resistance in Rainbow Trout are Achieved Using Low‐density SNP Panels: Evidence that Long‐range LD is a Major Contributing Factor. J. Anim. Breed. Genet. 135, 263–274. doi:10.1111/jbg.12335
VanRaden, P. M. (2008). Efficient Methods to Compute Genomic Predictions. J. Dairy Sci. 91, 4414–4423. doi:10.3168/jds.2007-0980
Varney, R. L., and Wilbur, A. E. (2020). Analysis of Genetic Variation and Inbreeding Among Three Lines of Hatchery-Reared Crassostrea virginica Broodstock. Aquaculture 527, 735452. doi:10.1016/j.aquaculture.2020.735452
Vera, M., Pardo, B. G., Cao, A., Vilas, R., Fernández, C., Blanco, A., et al. (2019). Signatures of Selection for Bonamiosis Resistance in European Flat Oyster (Ostrea edulis): New Genomic Tools for Breeding Programs and Management of Natural Resources. Evol. Appl. 12 (9), 1781–1796. doi:10.1111/eva.12832
Vu, S. V., Knibb, W., Nguyen, N. T. H., Vu, I. V., O'Connor, W., Dove, M., et al. (2020). First Breeding Program of the Portuguese Oyster Crassostrea angulata Demonstrated Significant Selection Response in Traits of Economic Importance. Aquaculture 518, 734664. doi:10.1016/j.aquaculture.2019.734664
Vu, S. V., Gondro, C., Nguyen, N. T. H., Gilmour, A. R., Tearle, R., Knibb, W., et al. (2021a). Prediction Accuracies of Genomic Selection for Nine Commercially Important Traits in the Portuguese Oyster (Crassostrea angulata) Using DArT-Seq Technology. Genes 12, 210. doi:10.3390/genes12020210
Vu, S. V., Knibb, W., Gondro, C., Subramanian, S., Nguyen, N. T. H., Alam, M., et al. (2021b). Genomic Prediction for Whole Weight, Body Shape, Meat Yield, and Color Traits in the Portuguese Oyster Crassostrea angulata. Front. Genet. 12, 661276. doi:10.3389/fgene.2021.661276
Walne, P. R., and Helm, M. M. (1979). Introduction of Crassostrea gigas into the United Kingdom. Cambridge: MIT Press, 83–105.
Wan, S., Li, Q., Yu, H., Liu, S., and Kong, L. (2020). Estimating Heritability for Meat Composition Traits in the Golden Shell Strain of Pacific Oyster (Crassostrea gigas). Aquaculture 516, 734532. doi:10.1016/j.aquaculture.2019.734532
Wang, J., Li, L., and Zhang, G. (2016). A High-Density SNP Genetic Linkage Map and QTL Analysis of Growth-Related Traits in a Hybrid Family of Oysters (Crassostrea gigas × Crassostrea angulata) Using Genotyping-By-Sequencing. G3 (Bethesda) 6, 1417–1426. doi:10.1534/g3.116.026971
Ward, R. D., Thompson, P. A., Appleyard, S. A., Swan, A. A., and Kube, P. D. (2005). Sustainable Genetic Improvement of Pacific Oysters in Tasmania and South Australia. FRDC Project 2000/206. CSIRO Marine and Atmospheric Research.
Xu, L., Li, Q., Yu, H., and Kong, L. (2017). Estimates of Heritability for Growth and Shell Color Traits and Their Genetic Correlations in the Black Shell Strain of Pacific Oyster Crassostrea gigas. Mar. Biotechnol. 19, 421–429. doi:10.1007/s10126-017-9772-6
Yang, B., Zhai, S., Zhang, F., Wang, H., Ren, L., Li, Y., et al. (2022). Genome-wide Association Study toward Efficient Selection Breeding of Resistance to Vibrio alginolyticus in Pacific Oyster, Crassostrea gigas. Aquaculture 548, 737592. doi:10.1016/j.aquaculture.2021.737592
Yin, X., Arias-Pérez, A., Kitapci, T. H., and Hedgecock, D. (2020). High-Density Linkage Maps Based on Genotyping-By-Sequencing (GBS) Confirm a Chromosome-Level Genome Assembly and Reveal Variation in Recombination Rate for the Pacific Oyster Crassostrea gigas. G3 (Bethesda) 10, 4691–4705. doi:10.1534/g3.120.401728
Yoshida, G. M., Bangera, R., Carvalheiro, R., Correa, K., Figueroa, R., Lhorente, J. P., et al. (2018). Genomic Prediction Accuracy for Resistance against Piscirickettsia salmonis in Farmed Rainbow Trout. G3 (Bethesda) 8, 719–726. doi:10.1534/g3.117.300499
Yoshida, G. M., Lhorente, J. P., Correa, K., Soto, J., Salas, D., and Yáñez, J. M. (2019). Genome-Wide Association Study and Cost-Efficient Genomic Predictions for Growth and Fillet Yield in Nile Tilapia (Oreochromis niloticus). G3 (Bethesda) 9, 2597–2607. doi:10.1534/g3.119.400116
Yuehuan, Z., Wu, X., Qin, Y., Xiao, S., Ma, H., Li, J., et al. (2017). Sustained Response to Selection of Growth Traits to the Third Generation for Two Strains of Kumamoto Oyster Crassostrea sikamea. J. Fish. Sci. China 24, 1161–1167. doi:10.3724/SP.J.1118.2017.16350
Zenger, K. R., Khatkar, M. S., Jones, D. B., Khalilisamani, N., Jerry, D. R., and Raadsma, H. W. (2019). Genomic Selection in Aquaculture: Application, Limitations and Opportunities with Special Reference to Marine Shrimp and Pearl Oysters. Front. Genet. 9, 693. doi:10.3389/fgene.2018.00693
Zhang, J., Li, Q., Xu, C., and Han, Z. (2019). Response to Selection for Growth in Three Selected Strains of the Pacific Oyster Crassostrea gigas. Aquaculture 503, 34–39. doi:10.1016/j.aquaculture.2018.12.076
Keywords: Ostrea edulis, oyster, GWAS, genomic selection, growth, aquaculture
Citation: Peñaloza C, Barria A, Papadopoulou A, Hooper C, Preston J, Green M, Helmer L, Kean-Hammerson J, Nascimento-Schulze JC, Minardi D, Gundappa MK, Macqueen DJ, Hamilton J, Houston RD and Bean TP (2022) Genome-Wide Association and Genomic Prediction of Growth Traits in the European Flat Oyster (Ostrea edulis). Front. Genet. 13:926638. doi: 10.3389/fgene.2022.926638
Received: 22 April 2022; Accepted: 17 June 2022;
Published: 15 July 2022.
Edited by:
Nguyen Hong Nguyen, University of the Sunshine Coast, AustraliaReviewed by:
Neil Thompson, Agricultural Research Service (USDA), United StatesPanya Sae-Lim, MOWI Genetics, Norway
Copyright © 2022 Peñaloza, Barria, Papadopoulou, Hooper, Preston, Green, Helmer, Kean-Hammerson, Nascimento-Schulze, Minardi, Gundappa, Macqueen, Hamilton, Houston and Bean. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Tim P. Bean, tim.bean@roslin.ed.ac.uk; Ross D. Houston, ross.houston@bmkgenetics.com
†These authors have contributed equally to this work