
Abstract
Background
Klebsiella aerogenes can cause ventilator-associated pneumonia by forming biofilms, and it is frequently associated with multidrug resistance. Phages are good antibiotic alternatives with unique advantages. There has been a lack of phage therapeutic explorations, kinetic studies, and interaction mechanism research targeting K. aerogenes.
Methods
Plaque assay, transmission electron microscopy and whole-genome sequencing were used to determine the biology, morphology, and genomic characteristics of the phage. A mouse pneumonia model was constructed by intratracheal/endobronchial delivery of K. aerogenes to assess the therapeutic effect of phage in vivo. Bioinformatics analysis and a prokaryotic protein expression system were used to predict and identify a novel capsule depolymerase. Confocal laser scanning microscopy, Galleria mellonella larvae infection models and other experiments were performed to clarify the function of the capsule depolymerase.
Results
A novel lytic phage (pK4-26) was isolated from hospital sewage. It was typical of the Podoviridae family and exhibited serotype specificity, high lytic activity, and high environmental adaptability. The whole genome is 40,234 bp in length and contains 49 coding domain sequences. Genomic data show that the phage does not carry antibiotic resistance, virulence, or lysogenic genes. The phage effectively lysed K. aerogenes in vivo, reducing mortality and alleviating pneumonia without promoting obvious side effects. A novel phage-derived depolymerase was predicted and proven to be able to digest the capsule, remove biofilms, reduce bacterial virulence, and sensitize the bacteria to serum killing.
Conclusions
The phage pK4-26 is a good antibiotic alternative and can effectively relieve pneumonia caused by multidrug-resistant K. aerogenes. It carries a depolymerase that removes biofilms, reduces virulence, and improves intrinsic immune sensitivity.
Similar content being viewed by others


Introduction
Klebsiella aerogenes, a common gram-negative, facultative anaerobic bacterium belonging to the family Enterobacteriaceae, has been reported to be an important opportunistic and multidrug resistant (MDR) pathogen in humans, particularly patients who are on mechanical ventilation [1]. From the early 1990s to 2003, K. aerogenes was the most clinically prevalent cause of nosocomial Enterobacter infections, and MDR K. aerogenes was an increasing cause of hospital-acquired infections [2]. In addition to its intrinsic resistance to ampicillin [3], K. aerogenes also has a broad ability to develop antibiotic resistance mechanisms [4, 5]. Moreover, there have been reports of carbapenem-resistant K. aerogenes with polymyxin resistance [6, 7], which may lead to the emergence of untreatable invasive infections.
Klebsiella aerogenes can form biofilms in endotracheal tubes and cause ventilator-associated pneumonia in mechanically ventilated patients [8, 9]. K. aerogenes strains with thicker capsules have higher antimicrobial resistance, stronger phagocytosis resistance, and lower serum sensitivity [10, 11]. There is a lack of effective and safe treatment options to control MDR K. aerogenes infection; therefore, developing alternatives to antibiotics to control colonization, remove established biofilms and reduce virulence is urgent and important.
As natural predators of bacteria, phages have the potential to address the absence of drugs in the post-antibiotic era [12]. Phages are host-specific and can effectively lyse bacteria. In recent years, phage therapy has been used to treat diseases caused by MDR pathogens, such as lung infections in transplant patients and bone and joint infections [13, 14]. In addition to the direct use of phages, other antimicrobial methods associated with phages include the use of endolysin and depolymerases [15]. Depolymerases can bind and specifically cleave polysaccharide-repeating units of host bacteria and destroy biofilms while antibiotics lack these abilities [16,17,18]. Compared with phages, the effect of on bacteria is more stable, consistent across strains with the same capsule type, and depolymerases have higher thermal stability, making them suitable for mass production [19]. In recent years, several studies have reported on phages that target K. aerogenes, but there has been a lack of phage therapeutic explorations, kinetic studies, and depolymerase mining.
In this study, a novel lytic phage (pK4-26) was isolated from hospital sewage, and its biology and genomic characteristics were determined. It can effectively lyse K. aerogenes in vivo, slow inflammation progression, and reduce mortality in mouse pneumonia models. Moreover, a novel phage-derived depolymerase (K4-26 dep protein) was predicted and indicated to be able to digest the capsule and degrade biofilms. After co-incubation with K4-26 dep protein, the bacteria showed lower virulence and higher innate immune sensitivity. This phage and the depolymerase it carries have the potential to be used in future antimicrobial therapy.
Methods
Bacterial strains, cells, and mice
All strains were isolated from clinical samples [20]. All strains were incubated in Luria–Bertani (LB) medium (Oxoid Ltd., UK) at 37 °C. An MDR strain (K. aerogenes 4-26) isolated from the endotracheal tube tip was used as a host for the isolation and proliferation of phage. Human lung carcinoma A549 cells were cultured in Dulbecco’s modified Eagle’s medium containing 10% fetal calf serum at 37 °C with 5% CO2. Male C57 BL/6J mice (7–8 weeks old) were purchased from the Beijing Vital River Laboratory Animal Technology Co., Ltd. (Beijing, China).
Isolation and biological characterization of phage pK4-26
Phage was isolated from organic sewage water in the Children’s Hospital Affiliated to Capital Institute of Pediatrics using the double-layer overlay technique. Phage titers were measured using a double agar overlay plaque assay. Different phage concentrations were mixed with log-phase host stains and co-culture for 3 h at 37 °C to measure the optimal multiplicity of infection (MOI). The host strain in log-phase was mixed with phage particles at the optimal MOI and incubated at 37 °C and the phage titers were calculated every 10 min over 150 min to generate a one-step growth curve. The temperature and pH stability of pK4-26 was determined by incubating phage preparations at different temperatures or pH values. After treatment, titers of residual phages were counted by double-layer plaque assay. The host ranges were examined using a standard spot test and an efficiency of plating (EOP) assay. The phage morphology was observed using transmission electron microscopy (TEM) operated at 80 kV.
Klebsiella aerogenes infection and phage therapy in mouse models
The minimum lethal dose (MLD)was determined, and the animal model was constructed according to previous reports [21, 22]. Briefly, the mice were randomly allocated into four groups: (1) the K. aerogenes group (n = 25), which received an intratracheal/endobronchial instillation of 2 × 107 CFU K. aerogenes (2 × MLD) and intranasal delivery of normal saline; (2) the K. aerogenes + pK4-26 group (n = 25), which received an intratracheal/endobronchial instillation of 2 × 107 CFU K. aerogenes and intranasal delivery of 2 × 109 PFU pK4-26; (3) the pK4-26 group (n = 25), which received an intratracheal/endobronchial instillation of normal saline and intranasal delivery of 2 × 109 PFU pK4-26; and (4) the paired group (n = 25), which received an intratracheal/endobronchial instillation of normal saline and intranasal delivery of normal saline. Three mice were euthanized to monitor bacterial loads and phage titers in the lungs at 2 h, 6 h, 12 h, 24 h, 48 h, 72 h and 7 days post infection. The lung tissues were also used for cytokine (IL-1β, IL-6, and TNF-α) level determination, and the lung, liver, kidney, and spleen tissues were used for hematoxylin–eosin staining at 24 h and 48 h post infection.
Genome sequencing and analysis
The genomic DNA of the phage was extracted using the phenol–chloroform method as previously described [23]. Whole-genome sequencing of DNA was performed on the Illumina HiSeq 2500 platform (Berry Genomics Corp., China). Potential coding domain sequences (CDS) were identified using PHASER [24], and putative homologies with predicted phage proteins were identified using BLASTP. A genomic map was constructed using the CGView Server. Phylogenetic trees based on amino acid sequences of the large terminase subunit, and the putative depolymerase gene sequences were constructed by MEGA 11, using the maximum likelihood method with 1000 bootstrap replicates. The resulting trees were plotted, annotated, and visualized using ggtree v3.2.1 [25]. Genome comparisons of pK4-26 and other phages were performed using Easyfig 2.2.5.
Protein domain and structure prediction
Protein domain prediction of the putative depolymerase was performed using BLASTP. The protein sequences of phages hosted by K. aerogenes were downloaded from the National Center for Biotechnology Information (NCBI) viral genome database and analyzed using PfamScan. Protein structure prediction of the putative depolymerase was performed using the Robetta server, and the predicted structure was visualized in PyMol2.4.
Expression and purification of putative depolymerase
The CDS (including stop codons) was inserted into the pET-28a expression vector via the BamHI and XhoI restriction sites. The primers and the cycling program are described in Additional file 3: Table S1. The recombinant plasmids were transformed into Escherichia coli BL21 cells and induced by adding 0.1 mM IPTG and incubating at 4 °C overnight. The resulting His-tagged protein was purified using Ni Sepharose 6FF (Solarbio, Beijing, China). The protein concentration was determined by the bicinchoninic acid method.
Determination of anti-capsule ability
Different concentrations of purified recombinant depolymerase were spotted into a lawn of the host strain to test its host range. TEM was used to observe the morphological changes in K. aerogenes 4-26 after incubation with 100 ng/μL of depolymerase solution at 37 °C for 6 h. The CPSs were extracted using a phenol-extraction method according to a previous method [26]. The capsular polysaccharides (CPSs) were incubated with different concentrations of the depolymerase for 12 h at 37 °C to verify their anticapsule ability. After incubation, the concentration of reducing sugars in each tube was determined using a 3,5-dinitrosalicylic acid (DNS) test kit (Solarbio, Beijing, China).
Formation, observation, and quantitation of biofilms
Bacteria cultured overnight were diluted 100-fold, transferred onto confocal dishes, and incubated at 37 °C for 24 h to form biofilms. After removing the supernatants and washing confocal dishes three times using PBS, different concentrations of depolymerase were added and incubated for 24 h at 37 °C. The formed biofilms were fixed using 4% glutaraldehyde for 3 h, stained with 50 μg/mL FITC-ConA for 20 min and then stained with 5 μg/mL PI for 15 min. The stained biofilms were visualized by confocal laser scanning microscopy (Leica TCS SP8 X, Germany).
Quantitation of biofilms was performed using a crystal violet staining method. The same bacteria as the above treatment were transferred onto 96-well flat-bottomed microtiter plates and incubated for 24 h at 37 °C. After removing the supernatants and washing the wells three times, 200 μL of PBS or different concentrations of depolymerase were added to the wells and incubated for 6, 12 and 24 h at 37 °C. After removing the supernatants and washing using with PBS, the wells were stained with a 0.75% crystal violet solution for 30 min and washed twice with purified water. The wells’ contents were solubilized in 200 µL of anhydrous ethanol for 30 min, and the optical density at 570 nm (OD570) was measured to quantify of the biofilms.
Bacterial virulence determination
A549 cells (4 × 105) were cultured in a 96-well plate and infected with different concentrations of K. aerogenes 4-26 for 3 h, which were pretreated with 100 ng/μL of depolymerase solution at 37 °C for 6 h or PBS. The number of viable cells was determined using the cell counting kit‐8 (CCK‐8) method (Beyotime Biotechnology Co., Shanghai, China). G. mellonella larvae infection models were also used to compare the virulence of bacteria. Briefly, different concentrations of bacterial or normal saline was injected into the last pair of pro-left legs. Then the larvae were incubated at 37 °C. Each group contained ten larvae. The time of death of each larva was recorded.
Serum killing assay
The bacteria were pretreated with depolymerase or PBS using the same method as above. Then, the bacteria were mixed with human serum from healthy volunteers and incubated at 37 °C for 6 h. The percent survival of bacteria was calculated based on viable counts relative to the initial inoculum. Three replicates were set up for each experimental condition.
Thermal and pH stability of depolymerase
The temperature and pH stability of depolymerase was determined by incubating depolymerase preparations (100 ng/μL) at different temperatures or pH values. After treatment, the preparations were incubated with equal amounts of CPS solution at 37 °C for 6 h. The concentrations of reducing sugars were determined using a DNS test kit. The percent survival of depolymerase was calculated based on the concentration of reducing sugars relative to the highest sugar concentration.
Statistical analysis
Data are presented as the mean ± SD and were analyzed with one-way ANOVA using SPSS 20.0 (IBM Corp., USA). Groups were compared with Kruskal–Wallis test for survival function. Differences were judged statistically significant at a P value of < 0.05.
Results
A novel lytic Podoviridae phage hosted by K. aerogenes was isolated
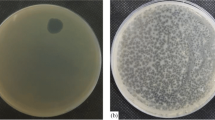
A bacterial strain (K. aerogenes 4-26) was isolated from the tip of a tracheal tube, and the strain showed multidrug resistance (Additional file 3: Table S2). A lytic phage named pK4-26 was isolated from hospital sewage using the strain as a host. The phage formed clear plaques with halos in double-layer agar plates, and the diameter of the plaques was 3.03 ± 0.34 mm (Fig. 1A). The phage was typical of the Podoviridae family, possessing an isometric head with a diameter of approximately 45 nm and short tails (Fig. 1B). It infected 6 of the 13 K. aerogenes strains in our laboratory (Table 1). The optimal MOI was 0.01, and a one-step growth curve showed that the latent period was 70 min, and the average burst size was 34.7 PFU/cell (Fig. 1C, D). The phage remained stable in environments below 50 °C and at pH values between 6 and 10 (Fig. 1E, F).

Morphology, biological characterization, and genomic features of phage pK4-26. A Phage plaques of pK4-26. B Phage morphology observed by TEM. C Optimal multiplicity of infection of phage. D One-step growth curve. E Thermal stability of pK4-26. F pH stability of pK4-26. G The genomic map of pK4-26. Values are expressed as mean ± SD (n = 3)
The genome of phage pK4-26 is 40,234 bp in length and there are 49 CDSs. The complete genome sequences of pK4-26 are most similar to Klebsiella phage KN3-1 (NC_048131.1), and the identity and query coverage between the two phages were 85.77% and 80%, respectively (Additional file 3: Table S3), which verified that pK4-26 is a novel phage. Furthermore, the genomic data show that pK4-26 carries no antibiotic resistance, virulence, or lysogenic genes (Fig. 1G, Additional file 3: Table S4). Therefore, this novel phage is suitable for phage therapy.
Phage pK4-26 can effectively lyse K. aerogenes in vivo and relieve pneumonia
To assess the therapeutic effect of pK4-26 in vivo, a mouse pneumonia model was constructed by intratracheal/endobronchial delivery of K. aerogenes 4-26 (Fig. 2A). We found that 2 × 107 CFU K. aerogenes 4-26 caused death in 70% of mice in 48 h to 7 days. However, intranasal administration of pK4-26 was effective in preventing mouse mortality (Fig. 2B). When the bacteria were delivered into the lungs of the mice, they grew rapidly and stabilized at 107–108 CFU/g. After phage treatment, the bacteria decreased rapidly by approximately 141-fold within 48 h while the phage titers increased 19-fold in 12 h. Phage titers in the phage therapy group stabilized at 107–108 PFU/g in 48 h while almost no phages were left in the group treated with pK4-26 alone (Fig. 2C, D), which suggests that pK4-26 can rapidly lyse bacteria and replicate itself in vivo.

Phage pK4-26 can effectively lyse K. aerogenes in vivo and relieve pneumonia. A Schematic diagram of the experimental procedure for phage therapy. B Survival of mice infected with K. aerogenes followed by phage therapy (Kruskal–Wallis’s test). C Changes in bacterial loads in the lungs were recorded for 48 h. D Changes in phage titers in the lungs were recorded for 48 h. E Anatomy, H&E staining (×100 and ×400 magnifications) of lungs sections from each group for 24 and 48 h. F Proinflammatory cytokine levels in the lungs at 24 and 48 h. G Histopathological scores of lung tissue sections. The histopathological score was related to the following distribution: 1, thickened alveolar walls; 2, edema; and 3, tissue parenchymatous lesions such as congestion and hemorrhage. Tissue sections were evaluated by two trained pathologists with the following scores: 0, no pathological lesion; 1, mild; 2, moderate; and 3, severe. Values are expressed as means ± the SD (n = 3 mice/group). Values are expressed as mean ± SD (n = 3). *P < 0.05; **P < 0.01; ****P < 0.0001 (one-way ANOVA)
Klebsiella aerogenes 4-26 caused lung congestion, distension, and hepatomegaly in mice within 24 h and lung necrosis by 48 h. Pathological staining showed collapsed alveolar structures, and diffuse neutrophil infiltration with scattered hemorrhaging and necrosis (Fig. 2E). Cytokines in the lungs were correspondingly elevated (Fig. 2F). Phage treatment prevented progressive necrosis in the lungs of mice and reduced lung cytokine levels; pathological staining also showed only slight alveolar wall thickening in the phage therapy group. More importantly, phage treatment did not cause damage to other organs, nor did it cause elevated cytokines (Fig. 2E, F, Additional file 1: Fig. S1). No resistant bacteria were detected during phage treatment (Additional file 2: Fig. S2).
Unique characteristics of the phage were discovered by comparative genomics
To clarify the unique characteristics of the phage, its genome was compared to those of other phages in public databases. The phylogenetic tree showed that phage pK4-26 was clustered with the genus Przondovirus and was on a separate branch (Fig. 3A). It has a similar genetic structure to those of phage KN3-1, phage KN1-1 (NC_048129.1), phage KN4-1 (NC_048130.1), and phage K5-2 (NC_047798.1) (Fig. 3B), and the unmatched regions in the pK4-26 genome are the first CDS (gp1). Very few possible proteins are homologous to this sequence in the NCBI database. Phylogenetic analysis (Fig. 3A) showed that the protein was divided into one branch with the hypothetical protein RU59_00038 (YP_009196380.1) of the Enterobacter phage phiEap-1, which infects Enterobacter aerogenes (K. aerogenes). None of the other homologous proteins on different branches were derived from phages that infect K. aerogenes, which suggests that the protein possibly has host specificity. Other homologous proteins were annotated as structural proteins or tailspike proteins, so the protein may be a structural phage protein. Moreover, the protein contains a Pectate_lyase_3 domain in the N-terminal region, which was preset as a parallel β-helix fold structure (Fig. 3D, E).

Bioinformatics analysis of phage pK4-26 and the putative depolymerase. A Phylogenetic tree based on amino acid sequences of the large terminase subunit using maximum likelihood method with 1000 bootstrap replicates. B Comparative genomic analysis. C Phylogenetic tree based on gene sequences of the putative depolymerase using maximum likelihood method with 1000 bootstrap replicates. D Protein structure prediction of the putative depolymerase. E Protein domain prediction of the putative depolymerase
A novel capsule depolymerase was found and can degrade biofilms
The putative gene (gp1) was cloned and expressed via a pET-28a expression system. The protein formed translucent plaques in a dose-dependent manner and had a similar host range to phage pK4-26. Interestingly, all hosts belonged to the PSgc4 type (Fig. 4A, Table 1). This protein made the capsule around the bacteria disappear, making the surface of the bacteria smooth to rough. It degraded the extracted CPS into reducing sugars (Fig. 4B, C) Thus, the protein was a novel capsule depolymerase. Furthermore, the depolymerase significantly reduced biofilms formed by K. aerogenes 4-26 in both time-dependent and dose-dependent manners, thereby reducing the number of bacteria attached to the biofilm (Fig. 4D, E). The results showed that 1 ng/μL K4-26 dep protein eliminated more than 80% of biofilms in 24 h.

K4-26 dep protein can digest capsules and degrade biofilms. A The translucent plaques formed by the pK4-26 and K4-26 dep protein. B Morphology of K. aerogenes 4-26 under TEM before and after treatment using depolymerase. C Color change in DNS reacted with K. aerogenes 4-26 CPS. D Fluorescence staining showed that the depolymerase can significantly reduce the amount of biofilm formed by K. aerogenes 4-26. E Crystal violet staining also proved the K4-26 dep protein can degrade biofilm formed by K. aerogenes 4-26. Values are expressed as mean ± SD (n ≥ 3). ****P < 0.0001 (one-way ANOVA)
K4-26 dep protein can reduce virulence, and improve sensitivity to innate immunity
The capsule is one of the sources of bacterial virulence, and its presence is associated with phagocytosis resistance and increased mortality in animal models. After incubation with the depolymerase, K. aerogenes 4-26 caused less cell death and lower death rate of in G. mellonella larvae (Fig. 5A, B). The depolymerase also improved the sensitivity of K. aerogenes 4-26 to innate immunity (Fig. 5C). Moreover, the K4-26 dep protein remained stable in environments below 60 °C (Fig. 5D) and at a pH between 6 and 10 (Fig. 5E), thus showing slightly higher thermal stability than phage pK4-26.

Functional analysis and characterization of K4-26 dep protein. A Survival curve of G. mellonella larvae under different treatments (Kruskal–Wallis’s test). B CCK-8 assay reflecting bacterial virulence under different treatments. C The survival of depolymerase-treated bacteria by serum killing. D Thermal stability of phage. E pH stability of phage. Values are expressed as mean ± SD (n ≥ 3). *P < 0.05; ****P < 0.0001 (one-way ANOVA)
Discussion
Klebsiella aerogenes is a member of the ESKAPE group, which is the most frequently isolated species in clinical infections, especially in immunocompromised patients and those hospitalized in intensive care units. It is frequently associated with a multidrug resistance phenotype [27]. Phages are a promising tool that can be used to address the absence of a drug in the post-antibiotic era. Phages have several advantages over antibiotics including strain specificity, lack of structural destruction of the intestinal microbiome and low development costs [28]. Therefore, finding new phages, establishing phage banks, and studying the interaction mechanisms between phages and host bacteria is of great significance.
In recent years, several studies have reported on phages that target K. aerogenes; however, these studies were limited to phage biology and genomic characteristics [23, 29,30,31]. There is a lack of kinetic research on clinical treatment with phages, and our understanding of the interaction mechanisms between bacteriophages and bacteria needs to be deepened. This study isolated a novel phage targeting K. aerogenes named pK4-26 from sewage. It is a new member of the Przondovirus genus. By closely monitoring bacterial and phage abundance over time in animal models, we demonstrated that the phage could replicate itself and lyse K. aerogenes in vivo, reducing mortality and alleviating inflammation.
Phages of the Podoviridae family often exhibit depolymerases. These enzymes can bind to the host bacteria’s CPS, exopolysaccharides, or lipopolysaccharides and specifically cleave polysaccharide repeat units [18]. Clear and round plaques with a halo are a sign that the phages carry depolymerases. The phage pK4-26 also formed plaques with a halo, so we speculated that pK4-26 may carry a depolymerase.
Phages of the Przondovirus genus have a similar genomic structure [32, 33]. In 2019, Pan et al. identified three Podoviruses infecting Klebsiella that encoded different capsule depolymerases digesting specific capsule types [32]. We compared the genetic structure of pK4-26 with those phages and found an unmatched region. Therefore, we suspected that the unmatched region (gp1) may encode a depolymerase.
Generally, depolymerases encoded by phages consist of two parts, one has depolymerization function; the other is annotated as a structural protein [34]. This protein was predicted to have a Pectate_lyase_3 domain. Phylogenetic analysis showed that the protein was divided into one branch with the hypothetical protein RU59_00038 (YP_009196380.1) of the Enterobacter phage phiEap-1, which showed highly similarity (identity: 97.95%, query coverage: 100%). None of the other homologous proteins were derived from phages that infect K. aerogenes, which suggests that the protein possibly has host specificity. Other homologous proteins were annotated as structural or tailspike proteins, so the protein may be a structural phage protein.
There are few publications relating to depolymerases of K. aerogenes phage. We downloaded 3879 protein sequences of 24 phages that infect K. aerogenes from the NCBI viral genome database and analyzed them to find special domains such as Pectate_lyase_3, Hydrolase, Endo-N-acetylneuraminidases, and Glycanase [35]. We discovered that there were few proteins with depolymerization functions (Table 2, Additional file 3: Table S5). The hypothetical protein RU59_00038 (YP_009196380.1) of Enterobacter phage phiEap-1 has a Pectate_lyase_3 domain. Klebsiella phage N1M2 has a putative structural lysozyme (QGH72084.1) containing a Glyco_hydro_19 domain that is a member of a family of glycoside hydrolases, and the protein may function as a lysozyme [36] or depolymerase [37]. The phage is known to be able to remove a preformed K. aerogenes biofilm [31]. vB_KaeM_KaOmega has a putative cell wall hydrolase containing a hydrolase_2 domain that may have lysozyme or depolymerase function [35]. The small number of these predicted proteins may be due to limited research on phages targeting K. aerogenes, highlighting our research’s importance.
The host range of Klebsiella phages and their extended depolymerases is thought to be associated with serotypes based on capsule features [38,39,40,41]. However, there is still no accepted standard for serotyping K. aerogenes. In 2019, Guo et al. established a molecular serotyping scheme based on surface polysaccharide antigens [42], and following the method, the hosts of pK4-26 and the K4-26 dep protein all belong to PSgc4. This means that they may be serotype specific. The PSgc4-type stains accounted for approximately 33% of clinical isolates, which means that pK4-26 and depolymerase may have a wider-spectrum killing power against clinical strains.
Furthermore, we discovered that the protein can efficiently degrade biofilms formed by K. aerogenes 4-26. However, when K4-26 dep protein was added in advance, biofilm formation was not inhibited, but enhanced (data not presented). This may be because the bacteria degraded the protein and used it to form thicker, tighter biofilms [43].
Considering that the capsule is an important virulence factor, we explored the effect of depolymerase on bacterial virulence and immune sensitivity. After treatment with the depolymerase, the virulence of K. aerogenes 4-26 to both animal and cell models was reduced. After losing the capsule, the immune evasion ability of K. aerogenes 4-26 was also reduced and its sensitivity to innate immunity improved. We then tested the temperature stability of the depolymerase and found that it had slightly higher temperature resistance than pK4-26.
Many studies have confirmed that phages have good efficacy and safety in treating pneumonia and intestinal inflammation caused by Klebsiella in animal models [44, 45]. In addition, the depolymerases derived from Klebsiella phages have been proven to have the ability to digest capsules, remove biofilms, and reduce bacterial virulence [46, 47], and they have potential uses such as reducing lethality, vaccine development and clinical strain typing [38, 48, 49]. More exploration of phage pK4-26 and its depolymerases is worth studying.
Conclusions
To our knowledge, this is the first phage kinetic study targeting K. aerogenes. Our data demonstrated that pK4-26 has good efficacy and safety for treating K. aerogenes infections in mouse models. It carries a depolymerase that removes biofilms, reduces virulence, and improves intrinsic immune sensitivity. Indeed, more preclinical studies and mechanistic studies are needed for clinical use.
Availability of data and materials
All data generated or analyzed during this study are included in this published article and its Additional files.
References
Alvarez-Marin R, Navarro-Amuedo D, Gasch-Blasi O, et al. A prospective, multicenter case control study of risk factors for acquisition and mortality in Enterobacter species bacteremia. J Infect. 2020;80(2):174–81.
De Oliveira D, Forde BM, Kidd TJ, et al. Antimicrobial resistance in ESKAPE pathogens. Clin Microbiol Rev. 2020;33(3):10–128.
Freney J, Husson MO, Gavini F, et al. Susceptibilities to antibiotics and antiseptics of new species of the family Enterobacteriaceae. Antimicrob Agents Chemother. 1988;32(6):873–6.
Davin-Regli A, Pages JM. Enterobacter aerogenes and Enterobacter cloacae; versatile bacterial pathogens confronting antibiotic treatment. Front Microbiol. 2015;6:392.
Passarelli-Araujo H, Palmeiro JK, Moharana KC, et al. Genomic analysis unveils important aspects of population structure, virulence, and antimicrobial resistance in Klebsiella aerogenes. FEBS J. 2019;286(19):3797–810.
Zhang X, Qu F, Jia W, et al. Polymyxin resistance in carbapenem-resistant Enterobacteriaceae isolates from patients without polymyxin exposure: a multicentre study in China. Int J Antimicrob Agents. 2021;57(2): 106262.
Da SK, de Almeida DSG, Moura Q, et al. Genetic diversity of virulent polymyxin-resistant Klebsiella aerogenes isolated from intensive care units. Antibiotics (Basel). 2022;11(8):1127.
Inglis TJ, Millar MR, Jones JG, et al. Tracheal tube biofilm as a source of bacterial colonization of the lung. J Clin Microbiol. 1989;27(9):2014–8.
Adair CG, Gorman SP, Feron BM, et al. Implications of endotracheal tube biofilm for ventilator-associated pneumonia. Intensive Care Med. 1999;25(10):1072–6.
Allen PM, Fisher D, Saunders JR, et al. The role of capsular polysaccharide K21b of Klebsiella and of the structurally related colanic-acid polysaccharide of Escherichia coli in resistance to phagocytosis and serum killing. J Med Microbiol. 1987;24(4):363–70.
Williams P, Lambert PA, Haigh CG, et al. The influence of the O and K antigens of Klebsiella aerogenes on surface hydrophobicity and susceptibility to phagocytosis and antimicrobial agents. J Med Microbiol. 1986;21(2):125–32.
Pires DP, Costa AR, Pinto G, et al. Current challenges and future opportunities of phage therapy. FEMS Microbiol Rev. 2020;44(6):684–700.
Van Nieuwenhuyse B, Van der Linden D, Chatzis O, et al. Bacteriophage-antibiotic combination therapy against extensively drug-resistant Pseudomonas aeruginosa infection to allow liver transplantation in a toddler. Nat Commun. 2022;13(1):5725.
Ferry T, Kolenda C, Laurent F, et al. Personalized bacteriophage therapy to treat pandrug-resistant spinal Pseudomonas aeruginosa infection. Nat Commun. 2022;13(1):4239.
Salmond GP, Fineran PC. A century of the phage: past, present and future. Nat Rev Microbiol. 2015;13(12):777–86.
Chegini Z, Khoshbayan A, Taati MM, et al. Bacteriophage therapy against Pseudomonas aeruginosa biofilms: a review. Ann Clin Microbiol Antimicrob. 2020;19(1):45.
Parasion S, Kwiatek M, Gryko R, et al. Bacteriophages as an alternative strategy for fighting biofilm development. Pol J Microbiol. 2014;63(2):137–45.
Knecht LE, Veljkovic M, Fieseler L. Diversity and function of phage encoded depolymerases. Front Microbiol. 2019;10:2949.
Fischetti VA. Exploiting what phage have evolved to control gram-positive pathogens. Bacteriophage. 2011;1(4):188–94.
Gan L, Yan C, Cui J, et al. Genetic diversity and pathogenic features in Klebsiella pneumoniae isolates from patients with pyogenic liver abscess and pneumonia. Microbiol Spectr. 2022;10(2): e264621.
Gan L, Fu H, Tian Z, et al. Bacteriophage effectively rescues pneumonia caused by prevalent multidrug-resistant Klebsiella pneumoniae in the early stage. Microbiol Spectr. 2022;10(5): e235822.
Anand T, Virmani N, Kumar S, et al. Phage therapy for treatment of virulent Klebsiella pneumoniae infection in a mouse model. J Glob Antimicrob Resist. 2020;21:34–41.
Zhao J, Zhang Z, Tian C, et al. Characterizing the biology of lytic bacteriophage vB_EaeM_phiEap-3 infecting multidrug-resistant Enterobacter aerogenes. Front Microbiol. 2019;10:420.
Arndt D, Grant JR, Marcu A, et al. PHASTER: a better, faster version of the PHAST phage search tool. Nucleic Acids Res. 2016;44(W1):W16–21.
Yu G, Lam TT, Zhu H, et al. Two methods for mapping and visualizing associated data on phylogeny using Ggtree. Mol Biol Evol. 2018;35(12):3041–3.
Campos MA, Vargas MA, Regueiro V, et al. Capsule polysaccharide mediates bacterial resistance to antimicrobial peptides. Infect Immun. 2004;72(12):7107–14.
Davin-Regli A, Lavigne JP, Pages JM. Enterobacter spp.: update on taxonomy, clinical aspects, and emerging antimicrobial resistance. Clin Microbiol Rev. 2019;32(4):e00002-19.
Marongiu L, Burkard M, Lauer UM, et al. Reassessment of historical clinical trials supports the effectiveness of phage therapy. Clin Microbiol Rev. 2022;35: e6222.
Mishra CK, Choi TJ, Kang SC. Isolation and characterization of a bacteriophage F20 virulent to Enterobacter aerogenes. J Gen Virol. 2012;93(Pt 10):2310–4.
Li E, Wei X, Ma Y, et al. Isolation and characterization of a bacteriophage phiEap-2 infecting multidrug resistant Enterobacter aerogenes. Sci Rep. 2016;6:28338.
Lewis R, Clooney AG, Stockdale SR, et al. Isolation of a novel jumbo bacteriophage effective against Klebsiella aerogenes. Front Med (Lausanne). 2020;7:67.
Pan YJ, Lin TL, Chen YY, et al. Identification of three podoviruses infecting Klebsiella encoding capsule depolymerases that digest specific capsular types. Microb Biotechnol. 2019;12(3):472–86.
Hsieh PF, Lin HH, Lin TL, et al. Two T7-like bacteriophages, K5-2 and K5-4, each encodes two capsule depolymerases: isolation and functional characterization. Sci Rep. 2017;7(1):4624.
Pires DP, Oliveira H, Melo LD, et al. Bacteriophage-encoded depolymerases: their diversity and biotechnological applications. Appl Microbiol Biotechnol. 2016;100(5):2141–51.
Latka A, Maciejewska B, Majkowska-Skrobek G, et al. Bacteriophage-encoded virion-associated enzymes to overcome the carbohydrate barriers during the infection process. Appl Microbiol Biotechnol. 2017;101(8):3103–19.
Ramesh N, Manohar P, Eniyan K, et al. A lysozyme murein hydrolase with broad-spectrum antibacterial activity from Enterobacter phage myPSH1140. Antimicrob Agents Chemother. 2022;66(9): e50622.
Hughes KA, Sutherland IW, Clark J, et al. Bacteriophage and associated polysaccharide depolymerases—novel tools for study of bacterial biofilms. J Appl Microbiol. 1998;85(3):583–90.
Lin TL, Hsieh PF, Huang YT, et al. Isolation of a bacteriophage and its depolymerase specific for K1 capsule of Klebsiella pneumoniae: implication in typing and treatment. J Infect Dis. 2014;210(11):1734–44.
Gorodnichev RB, Volozhantsev NV, Krasilnikova VM, et al. Novel Klebsiella pneumoniae K23-specific bacteriophages from different families: similarity of depolymerases and their therapeutic potential. Front Microbiol. 2021;12: 669618.
Domingo-Calap P, Beamud B, Mora-Quilis L, et al. Isolation and characterization of two Klebsiella pneumoniae phages encoding divergent depolymerases. Int J Mol Sci. 2020;21(9):3160.
Li M, Wang H, Chen L, et al. Identification of a phage-derived depolymerase specific for KL47 capsule of Klebsiella pneumoniae and its therapeutic potential in mice. Virol Sin. 2022;37(4):538–46.
Guo X, Wang M, Wang L, et al. Establishment of a molecular serotyping scheme and a multiplexed luminex-based array for Enterobacter aerogenes. Front Microbiol. 2018;9:501.
Secor PR, Sweere JM, Michaels LA, et al. Filamentous bacteriophage promote biofilm assembly and function. Cell Host Microbe. 2015;18(5):549–59.
Federici S, Kredo-Russo S, Valdes-Mas R, et al. Targeted suppression of human IBD-associated gut microbiota commensals by phage consortia for treatment of intestinal inflammation. Cell. 2022;185(16):2879–98.
Hesse S, Malachowa N, Porter AR, et al. Bacteriophage treatment rescues mice infected with multidrug-resistant Klebsiella pneumoniae ST258. MBio. 2021;12(1):e00034-21.
Li M, Li P, Chen L, et al. Identification of a phage-derived depolymerase specific for KL64 capsule of Klebsiella pneumoniae and its anti-biofilm effect. Virus Genes. 2021;57(5):434–42.
Pertics BZ, Cox A, Nyul A, et al. Isolation and characterization of a novel lytic bacteriophage against the K2 capsule-expressing hypervirulent Klebsiella pneumoniae strain 52145, and identification of its functional depolymerase. Microorganisms. 2021;9(3):650–69.
Hua Y, Wu Y, Guo M, et al. Characterization and functional studies of a novel depolymerase against K19-type Klebsiella pneumoniae. Front Microbiol. 2022;13:878800–9.
Li J, Sheng Y, Ma R, et al. Identification of a depolymerase specific for K64-serotype Klebsiella pneumoniae: potential applications in capsular typing and treatment. Antibiotics (Basel). 2021;10(2):144–61.
Acknowledgements
We thank American Journal Experts (http://bit.ly/AJE-HS), for editing the English text of a draft of this manuscript. We thank Institute of Biophysics (Chinese Academy of Sciences) for providing imaging and analysis services using TEM.
Funding
This work was supported by grants from the National Natural Science Foundation for Key Programs of China Grants (82130065), FENG foundation (FFBR 202103), National Natural Science Foundation of China (32170201), Beijing Natural Science Foundation (7222014), the Research Foundation of Capital Institute of Pediatrics (JCYJ-2023-08).
Author information
Authors and Affiliations
Contributions
JY (Corresponding Author): funding acquisition, resources, supervision, writing—review and editing, project administration. XC (First Author): conceptualization, methodology, formal analysis, validation, writing—original draft. BD (Co-first Author): methodology, resources, validation. JF: methodology, data curation, visualization. YF: methodology, resources. ZF, JC, JC, LG, TF: resources, methodology. ZT, RZ, CY, HZ, WX, ZX, ZY, ZD, ZL, YC: resources. GX (Co-corresponding Author): supervision, writing—review and editing. All authors approved the article.
Corresponding authors
Ethics declarations
Ethics approval and consent to participate
All animal experiments were approved by Capital Institute of Pediatrics Animal Care and Use Committee on the Ethics of Animal Experiments (Permission No. DWLL2021009) and were in accordance with the NIH Guidelines for the Care and Use of Laboratory Animals.
Consent for publication
Not applicable.
Competing interests
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Additional information
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary Information
Additional file 1: Figure S1.
H&E staining of liver, kidney, and spleen in different groups at 24 h and 48 h.
Additional file 2: Figure S2.
Phage susceptibility testing of bacteria. (A) Description of strain source. (B) Phage susceptibility testing of bacteria isolated from the lungs at 7 days in the treatment group.
Additional file 3: Table S1.
Primer sequences and PCR procedure used for prokaryotic expression. Table S2. The minimum inhibitory concentration (MIC) values of the bacteria mentioned. Table S3. Comparison of pK4-26 with other phage genome sequences in the NCBI database. Table S4. The putative open reading frames of phage pK4-26. Table S5. Protein domain prediction of Klebsiella aerogenes phage.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/. The Creative Commons Public Domain Dedication waiver (http://creativecommons.org/publicdomain/zero/1.0/) applies to the data made available in this article, unless otherwise stated in a credit line to the data.
About this article

Cite this article
Cui, X., Du, B., Feng, J. et al. A novel phage carrying capsule depolymerase effectively relieves pneumonia caused by multidrug-resistant Klebsiella aerogenes. J Biomed Sci 30, 75 (2023). https://doi.org/10.1186/s12929-023-00946-y
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/s12929-023-00946-y